XiaoMi-AI文件搜索系统
World File Search SystemTamir

MEVD – 301(A) 光电集成电路
MEVD – 301(A) 光电子集成电路 第一单元光波导理论:波导理论:一维平面波导、二维波导、超越方程、波导模式、模式截止条件。 第二单元光波导制造和特性:波导制造:沉积薄膜;真空沉积和溶液沉积、扩散波导、离子交换和离子注入波导、III-V 化合物半导体材料的外延生长、通过湿法和干法蚀刻技术塑造波导。波导特性:表面散射和吸收损耗、辐射和弯曲损耗、波导损耗测量、波导轮廓分析。 第三单元光耦合基础:横向耦合器。棱镜耦合器。光栅耦合器。光纤到波导耦合器。光波导之间的耦合。定向耦合器。定向耦合器的应用。单元 IV 导波调制器和开关:光调制器中使用的物理效应:电光效应、声光效应和磁光效应。波导调制器和开关。单元 V 半导体激光器和探测器:激光二极管。分布式反馈激光器。集成光学探测器。单元 VI 集成光学的最新进展:导波设备和应用的最新技术,例如光子开关、可调谐激光二极管、光学集成电路。文本/参考文献 1. T Tamir,《导波光电子学》,Springer-Verlag,1990 年 2. R Sysm 和 J Cozens,《光导波和设备》,McGraw-Hill,1993 年

生物ECE
在 ECE 中列出或交叉列出的课程:EE381V:基因组信号处理和数据科学 [Haris Vikalo] EE381V:分子编程 [David Soloveichik] EE380L:数据挖掘 [Joydeep Ghosh] EE380L1V:高级数据挖掘 [Joydeep Ghosh] EE371R:数字图像和视频处理 [Al Bovik] EE381K:数字视频 [Al Bovik] EE351M:数字信号处理 [Haris Vikalo] EE281K-6:估计理论 [Haris Vikalo] EE374K/385J-31:生物医学电子仪器设计 [John Pearce] EE338L/382V:模拟集成电路设计 [Nan Sun] EE381V:活动感知和识别 [Edison Thomaz] EE385J:生物医学成像模式 [Tom Yankeelov] EE385J:生物医学仪器 [Emily Porter] EE374L:生物医学工程的应用 [H. Grady Rylander III] EE385J-18:生物医学成像:信号/系统 [Tom Yankeelov] EE385J-32:生物医学工程项目 EE385V:脑机交互 [Jose del R. Millan] EE382V:现实世界中的复杂网络 EE381V:计算磁共振成像 [Jon Tamir] EE385V:神经工程 [Jose del R. Millan] EE381V:口语语言技术 [David Harwath]

2022 年 60 位人工智能领袖
特约作者:Abhishek Gupta、Agnis Stibe、Alex Guilbault、Alex Wang、Alf Rehn、Anand Ranganathan、Angeliki Dedopoulou、Anthony Mills、Arathi Sethumadhavan、Aruna Kolluru、Aruna Pattam、Badr Boussabat、Boyka Simeonova、Christian Rudin、Cynthia Rudin , 大卫·佩雷拉,迪玛·图尔钦、爱德华·皮泽-纳普、埃莉诺·沃森、艾玛·达克沃斯、恩里科·帕奈、伊娃·阿加帕基、乔治·帕努、古勒斯坦·拉德万、格特扬·博莱特、哈里·马曼加基斯、希曼什·阿拉哈巴迪、贾尔·里贝罗、雅罗斯拉夫·布拉哈、詹妮弗·维多利亚·斯库雷尔、若昂·阿泽维罗、约翰·阿布鲁多汤普森、乔恩·斯基尼尔·阿古斯特森、 Jordi Guitart、Joseph Yun、Jyotirmay Gadewadikar、Kashyap Kompella、Luca Sambucci、Marly Nika、Mathew Hughes、Mayte Hidalgo、Michael Wu、Mike Tamir、Nazar Zaki、Netanel Eliav、Prakash Natarajan、Andy Pardoe 教授、Eleni Mangina 教授、Raj Madan、理查德·本杰明斯、萨米兰·高什、谢林·马修、Siddha Ganju、Simon Greenman、Steve Shwartz、Sunil Kumar、Therese Svensson、Tirthajyoti Sarkar、Veena Calambur、Yannis Kalfoglou、Yasen Dimitrov 概念、制作和编辑:George Krasadakis 制作协助:Lucy Woodhead 原创视觉设计:Coy Chen

14-基因组编辑技术及其在动物育种中的可用性
针对农产品库存不足,无法满足世界人口快速增长带来的粮食需求、气候变化导致农场动物适应困难、各种广泛传播的疾病等问题,每天都有新的解决方案出现。科学家普遍认为,利用最近发展的基因组编辑技术可以解决这些问题。基因组编辑是通过核酸酶在基因组的指定位置创建位点特异性DNA双链断裂(DSB),然后通过同源重组(HDR)或非同源重组(NHEJ)方法之一修复双链断裂,从而产生基因组改变的方法。将这些方法与胚胎移植技术相结合并应用于动物养殖的主要目的是提高产量和品质,以及提高动物福利和抗病能力。本研究旨在阐明基因组编辑方法及其在畜牧业中的应用领域。
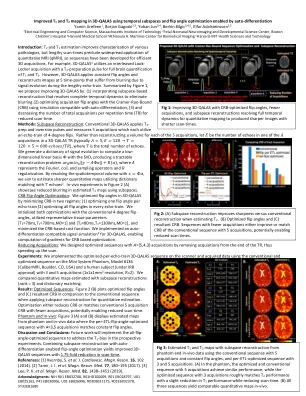
使用时间改进的 3D-QALAS 中的 T1 和 T2 映射...
简介:T 2 和 T 1 估计可改善各种病理的特征描述,但较长的扫描时间阻碍了定量 MRI (qMRI) 的广泛应用,因此已经开发了序列以实现高效的 3D 采集。例如,3D-QALAS 1 利用交错的 Look-Locker 采集和 T 2 准备脉冲来对 T 1 和 T 2 进行全脑量化。但是,3D-QALAS 应用恒定翻转角并在 5 个时间点重建图像,这些时间点由于冗长的回波序列期间的信号演变而出现模糊。总结图 1,我们建议通过以下方式改进 3D-QALAS:(1) 结合基于子空间的重建来解决完整的时间动态以消除模糊 (2) 使用与自动微分兼容的模拟通过 Cramer-Rao 界限 (CRB) 优化采集翻转角,(3) 并减少每重复时间 (TR) 的总采集次数以缩短扫描时间。方法:子空间重建:传统 3D-QALAS 应用 T 2 准备和反转脉冲并测量 5 次采集,每次采集都利用 4 度翻转的回声序列。不是为 5 次采集中的每次采集重建一个体积,而是让 𝐸 成为 3D-QALAS TR 中 𝐴 采集之一中的回声数量(通常 𝐴= 5,𝐸= 120 →𝑇= 120 × 5 = 600 𝑒𝑐ℎ𝑜𝑒𝑠/𝑇𝑅 ),其中 𝑇 是回声总数。我们生成一个信号演化字典,用 SVD 计算低维线性基 Φ,从而产生一个易于处理的重建问题 𝑎𝑟𝑔𝑚𝑖𝑛 𝛼 ‖𝑦−𝐴Φ𝛼‖ + 𝑅(𝛼) ,其中 𝐴 表示傅里叶、线圈和采样算子以及 𝑅 正则化。通过使用 𝑥= Φ𝛼 解析时空体积,我们旨在利用与 𝑇 回声 2 的字典匹配来估计更清晰的定量图。图 2 (A) 中的体内实验表明,使用子空间可以减少估计的 T 2 图中的模糊。 CRB 翻转角优化:我们通过最小化两种方式的 CRB 来优化 3D-QALAS 中的翻转角:(1) 优化每个回波序列的一个翻转角 (2) 优化每个回波序列中的所有翻转角。我们使用传统的 4 度翻转角初始化了这两种优化,利用了代表性组织参数 [T 2 =70ms、T 1 =700ms、M0=1] 和 [T 2 =80ms、T 1 =1300ms、M0=1],并最小化了基于 CRB 的成本函数。我们为 3D-QALAS 实现了自动微分兼容信号模拟 3,从而能够计算基于 CRB 的优化的梯度。减少采集:我们通过从 TR 末尾移除采集,设计了具有 A ={5,4,3} 采集的优化序列,从而加快了扫描速度。实验:我们在扫描仪上实施了针对每个回波序列进行优化的 3D-QALAS 序列,并使用 Mini System Phantom、型号 #136(CaliberMRI,美国科罗拉多州博尔德)和人类受试者(经 IRB 批准)上的常规和优化序列采集数据,进行了 3 次和 5 次采集(1x1x1mm3 分辨率,R=2)。我们比较了使用子空间重建(秩 = 3)和字典匹配估计的定量图。结果:优化序列:图 2(B)绘制了优化的翻转角和(C)与应用子空间重建进行定量估计时的传统序列相比的所得 CRB。优化可以减少 CRB 或者以更少的采集次数匹配传统的 5 次采集 CRB,从而有可能缩短扫描时间。模型和体内:图 3(A)和(B)显示了从模型和体内数据估计的图,其中每个 ETL 翻转角优化的序列(A=3,5 次采集)与恒定翻转角匹配。讨论和结论:未来的工作将实施全翻转角优化序列来解决未来实验中的 T 1 偏差。将子空间重建与自动微分启用的翻转角优化相结合,可获得改进的 3D-QALAS 序列,并将扫描时间缩短 1.75 倍。参考文献:[1] Kvernby, S. et al. J. Cardiovasc. Magn. Reson. 16 , 102 (2014)。[2] Tamir, JI 等人 Magn. Reson. Med. 77 , 180–195 (2017)。[3] Lee, PK 等人 Magn. Reson. Med. 82 , 1438–1451 (2019)。致谢:NIH R01 EB032708、R01HD100009、R01 EB028797、U01 EB025162、P41 EB030006、U01 EB026996、R03EB031175、R01EB032378、5T32EB1680