XiaoMi-AI文件搜索系统
World File Search Systemchromosomal

通过靶向有丝分裂激酶 PLK1 优先杀死四倍体结肠癌细胞
摘要背景/目的:染色体不稳定性是不同类型癌症(包括结直肠癌)进展的一个众所周知的因素。染色体不稳定性导致严重的核型重排和非整倍体。四倍体构成了致癌过程中多倍体/非整倍体级联的中间阶段,四倍体细胞对化疗特别有抵抗力。抑制有丝分裂蛋白 polo 样激酶 1 (PLK1) 是否会阻止四倍体结肠癌细胞的存活尚不清楚。方法:用 siPLK1 转染二倍体和四倍体细胞或用 PLK1 抑制剂 Bi2536 与纺锤体毒药联合处理。通过结晶紫染色和克隆形成测定评估细胞毒性。流式细胞术评估分析了许多细胞凋亡参数和细胞周期阶段。使用 CompuSyn 软件计算了 Bi2536 与紫杉醇、长春新碱或秋水仙碱之间的协同作用。结果:抑制或消除 PLK1 可阻止结肠癌细胞(特别是四倍体细胞)的存活。PLK 抑制引起的细胞死亡是由于有丝分裂滑移,随后激活了细胞凋亡的内在途径。我们进一步证明,用 PLK1 抑制剂和微管聚合抑制剂长春新碱或秋水仙碱(而不是微管解聚抑制剂紫杉醇)联合治疗四倍体结肠癌细胞会产生致命的协同效应。结论:PLK1 抑制与微管靶向化学物质相结合,可作为针对四倍体癌细胞的有效治疗策略。
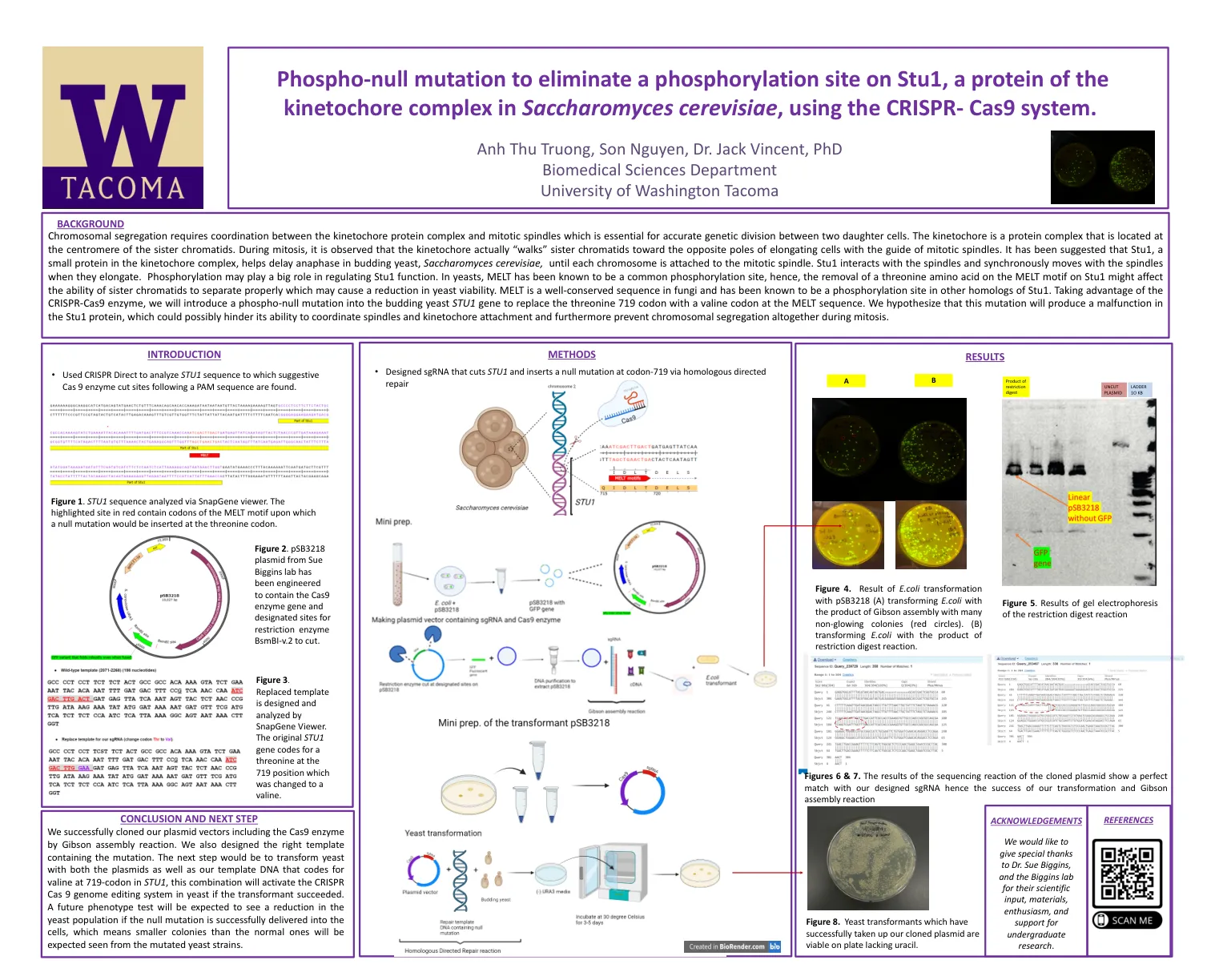
利用 CRISPR 技术进行磷酸化无效突变,消除酿酒酵母动粒复合体蛋白 Stu1 上的磷酸化位点
染色体分离需要动粒蛋白复合物和有丝分裂纺锤体的协调,这对于两个子细胞之间的准确遗传分裂至关重要。动粒是一种位于姊妹染色单体着丝粒的蛋白复合物。在有丝分裂过程中,可以观察到动粒实际上是在有丝分裂纺锤体的引导下将姊妹染色单体“引导”到伸长细胞的相反极点。有人提出,动粒复合物中的小蛋白 Stu1 有助于延迟芽殖酵母酿酒酵母的后期,直到每条染色体都附着在有丝分裂纺锤体上。Stu1 与纺锤体相互作用,并在纺锤体伸长时与其同步移动。磷酸化可能在调节 Stu1 功能方面发挥重要作用。在酵母中,MELT 是一种常见的磷酸化位点,因此,去除 Stu1 上 MELT 基序上的苏氨酸氨基酸可能会影响姐妹染色单体正确分离的能力,从而导致酵母活力下降。MELT 是真菌中保存良好的序列,并且已知是 Stu1 其他同源物中的磷酸化位点。利用 CRISPR-Cas9 酶,我们将在芽殖酵母 STU1 基因中引入磷酸化无效突变,以将 MELT 序列中的苏氨酸 719 密码子替换为缬氨酸密码子。我们假设这种突变会导致 Stu1 蛋白发生故障,这可能会阻碍其协调纺锤体和着丝粒附着的能力,并进一步阻止有丝分裂期间染色体分离。

针对去分化脂肪肉瘤中的 MDM2-p53 通路
去分化脂肪肉瘤 (DDLPS) 是一种恶性脂肪生成癌症,预后不良。DDLPS 肿瘤对化疗和放疗的敏感性较低,需要更有效的治疗方法。从遗传学上讲,DDLPS 的特点是肿瘤突变负担低,染色体结构异常频繁,包括 12q13-15 染色体区域和 MDM2 基因的扩增,这是 DDLPS 的定义特征。MDM2 蛋白是一种 E3 泛素连接酶,靶向肿瘤抑制因子 p53 进行蛋白酶体降解。人类恶性肿瘤中的 MDM2 扩增或过度表达与细胞周期进展和预后较差有关。因此,MDM2 - p53 相互作用作为 DDLPS 和其他恶性肿瘤的治疗靶点引起了人们的兴趣。MDM2 通过疏水蛋白相互作用与 p53 结合,这种相互作用很容易被合成类似物所利用。已经开发了多种药物,包括 Nutlins(如 RG7112)和小分子抑制剂(包括 SAR405838 和 HDM201)。临床前体外和动物模型已显示出抑制 MDM2 的良好效果,可导致 p53 强效再激活和癌细胞死亡。然而,多项早期临床试验未能证明抑制 MDM2 通路对 DDLPS 有益。耐药机制正在阐明,新型抑制剂和联合疗法目前正在研究中。本综述概述了针对 DDLPS 中 MDM2 的这些策略。

1 Genetic Testing: Whole Exome Sequencing and Whole Genome Sequencing
The human genome refers to an individual's complete set of DNA. The exome is a small section (1 to 2 percent) of the genome. It contains DNA sequences (exons) which provide instruction (coding) for making proteins, the building blocks of cells. Whole exome sequencing (WES) sequences only the coding region (1 to 2 percent) of an individual's genome. Whole exome sequencing can be used to identify variations in the protein-coding region of any gene rather than in only a select few genes. Because most known pathogenic variant(s) that cause disease occur in exons, WES is thought to be an efficient method to identify possible disease-causing pathogenic variant(s). Whole genome sequencing (WGS) sequences an individual's entire genome. It determines the order of all the nucleotides (the DNA building blocks) in an individual's DNA and can determine variations in any part of the genome. WES may potentially miss a pathogenic variant(s) in a non-coding region of the genome, therefore WGS may be used in selected cases if initial exome sequencing is not diagnostic. While WGS can accurately achieve copy number variation (CNV) detection, the use of chromosomal microarray analysis (CMA) continues to be the gold standard. Whole exome sequencing (WES) or whole genome sequencing (WGS) may be appropriate when there is no known cause of a patient's symptoms (e.g., prematurity, trauma, environmental, infectious, maternal immune disorder), clinical

CAS介导的植物染色体工程
最近的研究表明,不仅基因,而且整个染色体都可以使用定期间隔短的短膜重复序列(CRISPR)(CRISPR) - Crisper相关的蛋白9(Cas9)1 - 5进行设计。在植物育种中应用染色体重组的主要目标是操纵遗传交换6。在这里我们表明,使用染色体重组几乎可以在整个染色体中抑制减数分裂重组。我们能够诱导含有> 17 MB的染色体片段的可遗传反转,该片段包含着丝粒,并覆盖了拟南芥生态型Col-0的大部分染色体2。只有2和0.5 MB长的端粒末端保留在其原始原产中。在与生态型LER-1的杂交后代的单核苷酸多态性标志物分析中,我们检测到倒置的chrosome区域内的跨界群的大量降低,并伴随着交叉转移到远程端的末端。在反转中检测到的几种遗传交换都是源自双跨界的。这不仅表明可遗传的遗传交换可以通过间染色体配对来进行,而且还仅限于生存后代的产生。群集定期间隔短的短质体重复序列(CRISPR) - 基于危机相关的蛋白质(CAS)基因编辑已彻底改变了植物生物学和育种7。正在开发越来越多的工具来微调单基因和多个基因修饰8 - 10。能够改变染色体上基因的顺序也增加了一个新的特征控制水平:遗传联系的破裂11。为了将有吸引力的特征结合在单个培养基中,育种者通过减数分裂重组12之间的跨亲戚(CO)依赖于父母同种染色体之间的跨界(CO)12。众所周知,诸如倒置等染色体重排,通过抑制重排的区域13 - 18的CO来调节沿染色体的重组景观。例如,在果蝇中,所谓的平衡器染色体的特征是多种替代和其他重排,被广泛使用,导致抑制逆转杂合子中的减数分裂重组18。泛基因组的研究发现,自然染色体后序列在许多农作物物种中都是普遍存在的,并且在驯化4、19 - 24中发挥了重要作用。尽管它们看似善良,但反转也会导致积极影响,例如通过防止重组25来保护有利的等位基因组合。因此,CRISPR – CAS对染色体重排的有针对性诱导具有改变减数分裂重组模式的潜力。通过恢复1.1 MB大小的自然

使用 CEPT 混合物高效安全地进行 hPSC 单细胞克隆
CEPT 补充剂可促进健康单细胞克隆的建立。使用补充有 CEPT 混合物的培养基生成并接种微流体平台的克隆细胞系显示出与亲本系相似的增殖率和对单细胞解离的敏感性(图 3)。新的克隆系在培养中保持未分化状态,表达预期的多能性标记,并通过定向分化方法展示多能性。使用 CEPT 补充剂生成的克隆细胞系保持正常核型,在基因组癌症热点处未检测到染色体异常或 p53 突变。

DNA存储——从自然生物学到合成生物学
天然 DNA 存储可实现细胞分化、进化、我们孩子的成长,并控制我们所有的生态系统。在这里,我们讨论了 DNA 存储的基本方面和该领域的最新进展,特别强调了可以利用的自然过程和解决方案。我们指出了受自然启发的高效 DNA 和核苷酸存储的新方法。几年之内,基于 DNA 的信息存储可能会成为当前电子数据存储系统的有吸引力且自然的补充。我们讨论快速和定向访问(例如启动子、增强子等 DNA 元素)、调控信号和调制(例如 lncRNA)以及集成的高密度存储和处理模块(例如染色体区域)。有实用的 DNA 存储可用于生物技术和人类遗传学。我们将 DNA 存储作为合成生物学的一种方法(例如光控核苷酸加工酶)。DNA 和 RNA 的天然聚合物为直接存储操作(读入、读出、访问控制)提供了许多功能。内置的并行性(许多分子在许多地方同时工作)对于快速处理信息非常重要。使用染色体存储、核酸处理以及聚合物材料科学中的生物学概念(如酶、石墨烯、纳米纤维素直至 DNA 花边、DNA 线和基于 DNA 的适体场效应晶体管中的电子效应)将开辟新的应用,并随着时间的推移(几十年)逐渐取代越来越多领域的传统信息存储方法。

CALF的DNA依赖性RNA聚合酶的合成...
要描述的实验与组蛋白在核功能中的作用有关,特别强调了生物合成反应,这些反应通过引入乙酰基和甲基来改变组蛋白的结构。使用乙酸-C14和蛋氨酸 - 甲基-C'4在孤立的小牛胸腺核中研究了这些反应(参见参见参考文献1)作为前体,将它们的不合格与C14-赖氨酸和其他氨基酸的不合格进行比较,并测试普罗蛋白对不同组蛋白分数的合成的影响。将提供证据,以表明在细胞核中,组蛋白的乙酰化和甲基化很可能发生在多肽链完成后。尤其是乙酰化的组蛋白结构的这种修饰可能会影响组蛋白在体内抑制核糖核酸合成的能力。这种观点得到了以下发现的支持:当孤立的精氨酸组蛋白经过有限的乙酰化时,它们会因小牛胸腺核的DNA依赖性RNA聚合酶的RNA合成抑制剂而失去了许多有效性,因此它们的有效性很大。然而,这种修饰的组蛋白仍然是强烈的碱性蛋白质,它保留了与其得出的母体组蛋白相当的DNA的亲和力。这些发现介绍了组蛋白对核RNA的影响可能涉及的可能性不仅仅涉及对RNA合成的简单抑制,并且可能存在更微妙的机制,这些机制允许抑制和重新激活RNA沿染色体的RNA产生。在过去的几年中,对组蛋白作为染色体活性的调节剂的兴趣已大大提高,因为越来越多的实验证据已经积累了支持组蛋白的作用是抑制染色体

非同源末端连接是 CRISPR/Cas 的关键……
尽管成簇的规律间隔短回文重复序列 (CRISPR)/CRISPR 相关蛋白 (Cas) 介导的基因编辑已经彻底改变了生物学和植物育种,但大规模的可遗传植物染色体重组仍处于起步阶段。现在可以实现染色体内的重复和倒位,以及染色体之间的易位。随后,可以破坏或新建遗传连锁。此外,染色体上基因的顺序也可以改变。虽然自然染色体重组在减数分裂过程中通过同源重组发生,但 CRISPR/Cas 介导的染色体重排最好通过利用体细胞中的非同源末端连接 (NHEJ) 途径获得。NHEJ 可细分为经典 (cNHEJ) 和替代 NHEJ (aNHEJ) 途径,它们部分地以拮抗方式运作。 cNHEJ 通路不仅可以保护断裂的 DNA 末端免于降解,还可以抑制先前未连接的断裂末端的连接。因此,在没有 cNHEJ 的情况下,可以获得更多的倒位或易位,这可以归因于无限制地使用 aNHEJ 通路进行双链断裂 (DSB) 修复。与倒位或易位相反,短串联重复可以通过 Cas9 切口酶由成对的单链断裂产生。有趣的是,cNHEJ 通路对于这些类型的重复至关重要,而 aNHEJ 则是补丁插入所必需的,补丁插入也可以在 DSB 修复期间形成。由于染色体工程不仅在模式植物拟南芥 (Arabidopsis thaliana) 中实现,而且在作物玉米 (Zea mays) 中也实现,我们预计这项技术将很快改变育种过程。