XiaoMi-AI文件搜索系统
World File Search Systemspeed

航空航天和国防对速度的需求
大多数组件的生产都需要大量的非经常性成本 (NRC)。例如,工具、研发、资本设备、工程和测试等一次性费用可能占生产给定零件、模块或系统成本的大部分,从而限制了可以竞争生产它们的潜在供应商数量。对于某些商品,这可能会导致缺乏竞争,使供应商在与 OEM 打交道时拥有定价权,并可能使他们在实现进度和质量目标方面反应迟钝。其他商品需要相互渗透的关系和责任,需要更多的整理、更多的数据交换以及对权衡或需求变化影响的联合评估:所谓的“风险共担伙伴关系”模式仍需进行微调,以确保总承包商与具有广泛责任的设计和建造供应商之间的关系协调一致且有效。

基因驱动速度
利用遗传系统将所需性状与染色体或遗传因素联系起来,具有正交易偏见(即,> 50%)可以追溯到Serebrovski 1染色体易位的潜在用途,Serebrovski 1的潜在用途是curtis在1960年代在1960年代进一步概括和表达的,以遍及整个目标人群。这些所谓的基因驱动系统或自私基因3本质上很丰富。驾驶元素会偏向于性染色体或常染色体(减数分裂驱动)4 - 11或仅仅是自己,这是由可转移元素的不同家族12,13(例如,p -p- p- p- p- p- p- p- p- p- p- p- 14 - 16中的元素或人类中的人类17)17)。这种超级孟德尔遗传实体与动植物中的基因组结构的演变有关17 - 21。基因驱动器可以根据人群的易于传播的方式将基因驱动器广泛分为两个主要类别。高阈值驱动器,例如柯蒂斯认为2的相互染色体易位,需要许多人(例如,超过本地居民的数量)接管人口

解释“曲速行动”
• 8 月 11 日:美国卫生与公众服务部宣布拨款高达 15 亿美元,用于支持 Moderna 公司研究性候选疫苗的大规模生产和交付。根据协议条款,美国政府将拥有由此产生的 1 亿剂疫苗,并可选择购买更多疫苗。这种名为 mRNA-1273 的疫苗由 Moderna 公司和美国国立卫生研究院下属的国家过敏和传染病研究所 (NIAID) 的科学家共同开发。NIAID 继续支持该疫苗的开发,包括非临床研究和临床试验。此外,BARDA 还支持了该疫苗的 2/3 期临床试验、疫苗生产规模扩大和其他开发活动。3 期临床试验于 7 月 27 日开始,是美国政府资助的首个 COVID-19 疫苗 3 期临床试验。


航空航天和国防对速度的需求
大多数组件的生产都需要大量的非经常性成本 (NRC)。例如,工具、研发、资本设备、工程和测试等一次性费用可能占生产特定零件、模块或系统成本的大部分,从而限制了可以竞争生产这些产品的潜在供应商数量。对于某些商品而言,这可能会导致缺乏竞争,使供应商在与 OEM 打交道时拥有定价权,并可能使其在实现进度和质量目标方面反应迟钝。其他商品需要相互渗透的关系和责任,需要更多的整理、更多的数据交换以及对权衡或需求变化的影响的联合评估:所谓的“风险共担伙伴关系”模式仍需进行微调,以确保总承包商与具有广泛责任的设计和建造供应商之间的关系协调一致且有效。

生活速度。 - ecobat电池
永远不要停止重新思考。时代不断变化 - 在我们的行业中,甚至还有一个更重要的常数:Exide Technologies对创新和推动事物向前发展的愿望。,我们用高级产品证明了这一点。我们为各种动力总成技术提供了最大的电池范围之一。基于原始设备业务的专业知识,我们处于提供最先进的解决方案的最前沿。我们产品中无与伦比的性能使我们可以强调作为领先的OE品牌的可靠性。外观还提供了一套专业配件,允许讲习班为客户提供最高水平的服务。
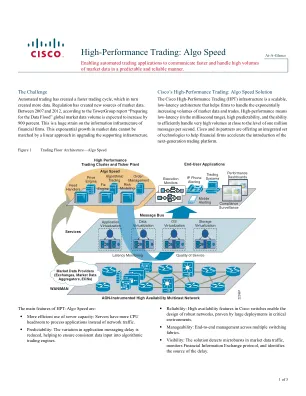
高性能交易:算法速度
•有助于准备新的法规:Reg NMS和MiFID等监管更改将生成更多报价,订购和取消/替换消息,因为股权公司适应了更多的电子业务流程。在美国,Subpenny-Presing规则还将增加对支持基础设施的需求。 MiFID还将导致更高的数据量,因为将其内部化交易的投资银行以电子方式发布。 交易应用程序的加速有助于确保最佳执行要求并提高交易竞争优势。在美国,Subpenny-Presing规则还将增加对支持基础设施的需求。MiFID还将导致更高的数据量,因为将其内部化交易的投资银行以电子方式发布。交易应用程序的加速有助于确保最佳执行要求并提高交易竞争优势。

利用人工智能加速流程并推动洞察
智能数据处理:利用 AI 模型从任何文档中获取文本和数据,无论其格式或内容如何。这些模型无需人工干预即可识别和提取相关信息,无论文档是贷款申请、医疗表格、法律合同还是其他内容。全面了解团队绩效,发现改进机会并确定可行的策略。
