XiaoMi-AI文件搜索系统
World File Search Systemsyngeneic

用于开发卵巢癌联合疗法的基因定义的同基因类器官平台
靶向疗法和免疫疗法并行开发,通常使用不同的实验系统。即使在今天,靶向药物通常也会针对癌细胞系/细胞来源的异种移植 (CDX)、患者来源的异种移植 (PDX) 和/或最近的人类肿瘤球体/类器官进行测试。这些模型的优势包括其人类起源、相关的突变/表观遗传事件以及保留一定程度的肿瘤异质性。然而,这样的系统无法评估抗肿瘤免疫反应。PDX 已在“人源化”小鼠中建立,但约 30% 的人/小鼠生长因子、细胞因子和趋化因子无法与其他物种中的同源受体相互作用,从而对“人源化”施加了内在限制 (Walsh 等人,2017)。相比之下,免疫疗法主要针对同源小鼠肿瘤进行测试 (Mosely 等人,2017)。这些模型(例如 B16、CT26 和 MC38)主要由致癌物引起,来源于未知、无关或并非最相关的细胞,并且通常缺乏相应人类疾病中发现的关键致病突变。一些靶向药物/免疫疗法已在基因工程小鼠模型(GEMM)中进行了评估,这些模型旨在携带与疾病相关的基因异常并具有完整的免疫系统(Kersten et al., 2017)。通常,对于给定的恶性肿瘤仅会产生少数突变组合,这限制了可分析的人类疾病的多样性。大多数 GEMM 还会同时将癌症相关缺陷引入目标组织的所有上皮细胞。相比之下,现实世界的肿瘤以克隆方式起始,并在以正常细胞为主的海洋中扩增和进展。已经生成了一系列可移植的 GEMM 衍生黑色素瘤模型(Yum/Yummer)(Meeth 等,2016),但这些都是基于相同的躯干突变,具有有限的遗传多样性。

抗 VEGF 疗法可改善小鼠同基因胶质母细胞瘤模型中的 EGFR-vIII-CAR-T 细胞递送和疗效
摘要 嵌合抗原受体 (CAR)-T 细胞彻底改变了多种血液系统恶性肿瘤的治疗方法,但在患有胶质母细胞瘤 (GBM) 或其他实体瘤的患者中显示出有限的疗效。这可能主要是由于免疫抑制肿瘤微环境 (TME) 损害了 CAR-T 细胞的输送和抗肿瘤活性。我们之前表明,阻断血管内皮生长因子 (VEGF) 信号传导可以使小鼠和人类肿瘤(包括 GBM、乳腺癌、肝癌和直肠癌)中的肿瘤血管正常化。此外,我们证明血管正常化可以改善 CD8+ T 细胞的输送和小鼠乳腺癌模型中免疫治疗的疗效。事实上,美国 FDA(食品和药物管理局)在过去 3 年中已经批准了七种不同的抗 VEGF 药物和免疫检查点阻滞剂组合,用于治疗肝癌、肾癌、肺癌和子宫内膜癌。在这里,我们测试了以下假设:抗 VEGF 疗法可以改善 CAR-T 细胞在携带原位 GBM 肿瘤的免疫活性小鼠中的输送和疗效。我们设计了两种同源小鼠 GBM 细胞系 (CT2A 和 GSC005) 以表达 EGFRvIII(人类 GBM 中最常见的新抗原之一),并设计了 CAR T 细胞以识别 EGFRvIII。我们发现,与单独使用 EGFRvIII-CAR-T 细胞疗法相比,使用抗小鼠 VEGF 抗体 (B20) 治疗可改善 CAR-T 细胞在整个 GBM TME 中的浸润和分布,延缓肿瘤生长,并延长 GBM 携带小鼠的生存期。我们的研究结果为临床评估抗 VEGF 药物与 CAR T 细胞对 GBM 患者的疗效提供了令人信服的数据和理论依据。

在合成小鼠肿瘤模型中具有低免疫细胞浸润的低氧CAIX阳性肿瘤区域的定量成像。 Boreel,D.F。; Span,P.N。;
摘要:氧与氧气消耗量增加的有限扩散导致大多数固体恶性肿瘤的慢性缺氧。已知这种氧气的稀缺性会诱导辐射势并导致免疫抑制的微环境。碳酸酐酶IX(CAIX)是一种酶,充当低氧细胞中酸性输出的催化剂,是慢性缺氧的内源性生物标志物。这项研究的目的是开发一种放射标记的抗体,该抗体识别出鼠类caix可视化慢性肿瘤模型中的慢性缺氧,并研究这些低氧区域中的免疫细胞群体。将一种抗MCACIS抗体(MSC3)偶联到二乙基三环乙酸乙酸(DTPA),并用依赖二醇标记为111(111英寸)。使用流式细胞仪确定鼠肿瘤细胞上的CAIX表达,并在竞争性结合测定中分析了[111 in] In-MSC3的体外亲和力。进行了体内生物分布研究,以确定体内放射性分布。CAIX +肿瘤分数通过MCAIX微光谱/CT确定,并使用免疫组织化学和自身自显影分析肿瘤微环境。我们表明,[111 in] In-MSC3在体外与表达Caix(Caix +)鼠细胞结合,并在体内积聚在Caix +地区。我们优化了[111 in] In-MSC3用于临床前成像的使用,以便可以将其应用于合成小鼠模型中,并表明我们可以通过Vivo McAix Micropect/CT进行定量区分具有不同CAIX +分数的肿瘤模型。对肿瘤微环境的分析确定这些Caix +区域被免疫细胞浸润较少。这些数据共同表明,McAix Microspect/CT是一种敏感技术,可视化缺氧的Caix +肿瘤区域,在合成小鼠模型中表现出降低免疫细胞的浸润。将来,该技术可能会在针对缺氧或减少缺氧治疗之前或期间可视化CAIX表达。因此,它将有助于优化翻译相关的合成小鼠肿瘤模型中的免疫和放射疗法功效。关键词:碳酸酐酶IX,缺氧,动物成像,免疫学,肿瘤微环境■简介

NCI药物开发研讨会:如何推进...
•syngeneic(免疫能力) - 肿瘤和宿主是近交菌株•同种异体(可变) - 相同的物种肿瘤和宿主并非完全杂交•异,肿瘤(免疫受损) - 肿瘤和宿主来自不同的物种
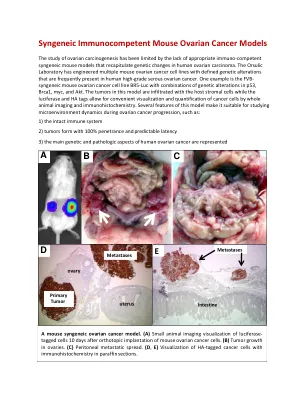
同基因小鼠卵巢癌细胞系-...
同基因免疫功能小鼠卵巢癌模型 由于缺乏合适的免疫功能同基因小鼠模型来重现人类卵巢癌的基因变化,卵巢癌的研究受到了限制。Orsulic 实验室已设计出多种小鼠卵巢癌细胞系,这些细胞系具有明确的基因改变,这些改变经常出现在人类高级别浆液性卵巢癌中。一个例子是 FVB 同基因小鼠卵巢癌细胞系 BR5-Luc,其具有 p53、Brca1、myc 和 Akt 基因改变的组合。该模型中的肿瘤浸润有宿主基质细胞,而荧光素酶和 HA 标签可通过整体动物成像和免疫组织化学方便地可视化和量化癌细胞。该模型的几个特点使其适合研究卵巢癌进展过程中的微环境动态,例如:

JAB-X1800:一种针对 CD73 的强效免疫刺激抗体-药物偶联物 (iADC)
图 7. JAB-X1800 的体内抗肿瘤活性和耐受性。A. JAB-X1800 在表达人 CD73 转基因 (hCD73-MC38) 的 MC38 结肠同种模型中的抗肿瘤活性。B. 用载体或 JAB-X1800 治疗原始肿瘤植入 (A) 的存活小鼠再次接受 hCD73-MC38 肿瘤细胞攻击,结果显示 JAB-X1800 治疗小鼠再次攻击后的肿瘤生长受到抑制。C. JAB-X1800 在 CD73 阴性 MC38 结肠同种模型中的抗肿瘤活性,该模型不表达人 CD73。D. JAB-X1800 在 CD73 阳性乳腺癌异种移植模型 (MDA-MB-231) 中的抗肿瘤活性。 E. JAB-X1800 在 CD73 阳性非小细胞肺癌异种移植模型 (NCI- H1975) 中的抗肿瘤活性。体重显示动物耐受性。每组 4-8 只小鼠。有效载荷是游离 STING 激动剂,CD73 抗体是未结合的抗人 CD73 抗体。同种型 ADC 是与同种型抗体结合的 STING 激动剂。

摘要:S154标题:第一类MTHFD1抑制剂...
(c)。因此,我们表明TH9619显着提高了人类肿瘤异种移植模型(OSU-CLL,MEC-1)(D)的小鼠的存活,并且完全消除了已经建立的原发性肿瘤(CLL细胞的S.C植入)。在高度侵略性的RS合成模型中也证明了TH9619的这种令人印象深刻的活动,

FAK抑制与RAF/MEK夹具Avutometinib相结合,克服了对BRAF和MEK
虽然BRAFV600E黑色素瘤Yumm 1.7合成肿瘤未能对ICI疗法做出反应,但添加了Avutometinib±Faki抑制了肿瘤的生长。我们观察到,用ICI + Avutometinib治疗的肿瘤最终产生了抗药性并逃脱了生长抑制,但是用ICI +合并的Avutometinib和Faki治疗的肿瘤表现出耐用的治疗反应,通常具有完全肿瘤的消退。

ENPP1 抑制剂 TXN10128
竞争力 • TXN10128 是一种强效且选择性的 ENPP1 抑制剂,可在 3D 共培养条件下发挥免疫反应。 • TXN10128 与抗 PD-L1 抗体一起增强了协同肿瘤生长抑制作用,并在 MC38 同基因小鼠模型中具有良好的 TIL 特征。 • TXN10128 具有良好的药物相似性和 PK 特征。 • TXN10128 是作为现有免疫疗法的联合用药进行临床研究的合适候选药物。 • TXN10128 的临床前研究将于 2022 年第四季度完成,第 1 阶段临床试验将于 2023 年开始。

有丝分裂MTH1抑制剂TH1579通过CGAS刺激途径诱导PD-L1的表达和炎症反应
有丝分裂MTH1抑制剂TH1579是一种双重抑制剂,可抑制有丝分裂和掺入氧化DNA损伤并导致特定于癌症的细胞死亡。通过CGAS刺激途径,DNA损害剂会增强对免疫检查点抑制剂(ICI)处理的反应。这项研究研究了TH1579是否可以通过其免疫调节特性改善免疫检查点阻滞的效率。用有丝分裂的MTH1I TH1579处理了各种人和鼠类癌细胞系,并通过流量细胞仪和实时QPCR分析了PD-L1和T细胞与燃料相关的趋化因子的表达。合成小鼠模型,以检查TH1579和PD-L1阻滞的综合作用。在我们的研究中,我们发现TH1579在人类癌细胞系中的蛋白质和mRNA水平上都上调了PD-L1的表达。但是,在鼠细胞系中,增加的增加不太明显。在合成小鼠黑色素瘤模型中的一个体内实验表明,与媒介物或atezolizumab单疗法相比,Th1579的治疗显着提高了Atezolizumab(一种抗PD-L1抗体)Atezolizumab(一种抗PD-L1抗体)。此外,Th1579表现出免疫调节特性,以CGAS丁字途径依赖性方式升高了细胞因子,例如IFN-β和包括CCL5和CXCL10在内的趋化因子和趋化因子。总而言之,TH1579具有通过调节免疫检查点相关蛋白和途径来改善ICI处理的潜力。