XiaoMi-AI文件搜索系统
World File Search Systemtemp

开发支持数字工程工件...
Edaptive Computing 的智能文档 Edaptive 正在为 MBTEMP 增强的成熟模型支持的文字处理和文档内容管理系统已得到演示。该解决方案将内容模块(例如系统描述)存储在后端数据库中,以便可以以版本控制的方式在多个文档中使用它们。该解决方案集成到 Microsoft Word 中,以便文档开发人员可以继续在其本机环境中工作。该系统目前正在增强,以将 IDSK 表从后端 SQL 数据库拉入 Microsoft Word 开发的 TEMP。未来的迭代将允许从一组模板直接在 Web 应用程序中完整生成 IDSK 数据库,而无需用户具备任何 SQL 知识。

蓝色印章™
标准和认证:• 经 NSF International 认证,符合 NSF/ANSI 标准 14 和 61,可用于饮用水、排水、废水、通风和下水道应用 • 经 IAPMO 认证,符合 IAPMO PS 36-2014e1 和最新版《统一管道规范》的适用部分 • FBC™ 系统兼容表示该产品已经过测试并持续受到监控,以确保其与 FlowGuard Gold®、BlazeMaster® 和 Corzan® 管道系统以及采用 TempRite® 技术制造的产品具有化学兼容性。FBC 系统兼容徽标、FBC™、FlowGuard Gold®、BlazeMaster®、Corzan® 和 Temp Rite® 是 Lubrizol Advanced Materials, Inc. 或其附属公司的商标。• 可根据 LEED®(能源与环境设计先锋)、IEQ Credit 4.1 获得积分

2023 年 3 月 10 日俄勒冈州经济分析办公室 Josh ...
• 注意此数据使用 IPUMS 一致定义的行业和职业代码,但仍存在潜在问题 • 这就是为什么建筑业整体生产率没有提高的原因吗?同样的工作量需要更多的工人? • 将手工工作外包给分包商或临时工,而这些分包商或临时工在其他地方被归类?(家庭调查结果与个体经营和受薪工作类似) • 项目组合发生变化? • 监管/合规性增强? • 其他行业特定问题?

C-130J - 主任操作测试和评估
活动 • 空军正在纠正开发和运行测试中发现的缺陷,并在 Block Upgrade 7.0 中添加新功能。Block Upgrade 7.0 已经历了大约一年的计划延迟;FDE 现在预计将在 2013 财年第一季度进行。 • DOT&E 于 2010 年 11 月批准了 C-130J 测试和评估总体计划 (TEMP),其中包括 Block Upgrade 7.0 和 SKE 软件增强测试。 • 空军根据 DOT&E 批准的测试计划于 2011 年 2 月和 3 月对 SKE 软件增强进行了 FOT&E。DOT&E 将在 2012 财年发布有关 SKE FOT&E 的报告。 • 空军于 2011 年 4 月完成了数据传输和诊断系统 (DTADS) 的开发测试和评估 (DT&E)。DTADS 将取代当前的
原始卖出签名和替代药物研究所( div>
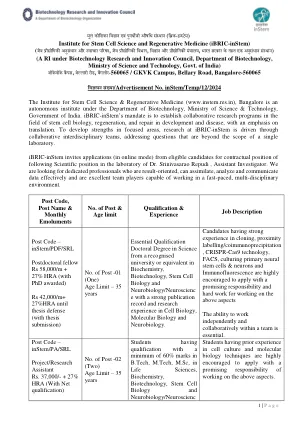
विजविजपनसंख /广告号< /div>Instem/Temp/12/2024班加罗尔干细胞科学与再生医学研究所(www.instem.res.in)是印度政府科学技术部生物技术部的自治研究所。ibric-Instem的任务是在干细胞生物学,再生和开发和疾病的修复领域建立合作研究计划,重点是翻译。为了在重点领域发展优势,IBRIC-INSTEM的研究是通过协作跨学科团队驱动的,解决了超出一个实验室范围的问题。IBRIC-INSTEM邀请合格候选人的申请(以在线模式),以在助理调查员Srinivasarao Recudi博士实验室中遵循科学地位的合同地位。我们正在寻找专门的专业人士,这些专业人员以结果为导向,可以有效地吸收,分析和传达数据,并且是能够在快速节奏的多学科环境中工作的优秀团队参与者。


供应实验室的招标文件...
参数电导率/tds/℃射程符号(0〜2×105)US/cm1.0.00μs/cm- 100ms/cm/cm TDS -0-1000 mg/ltemp -(0.0〜60.0.0)℃(0.0〜60.0)℃精度±1.0%fs±1.0%fs ±1位温度。补偿(5〜35)℃,手动(0.0〜40.0)℃,自动/手动功率220V/110V尺寸/N.W。300×200×90 mm / 1.5kg运输尺寸 / G.W。330×260×280 mm / 2kg < / div>

IQ电池10TIQ电池10T
1。从交替的电流到电流的交替电流到25°C的标称功率的50%(生命开始时)。实际回报产量可能会根据环境温度,负载模式和其他外部因素而有所不同。2。在日常操作的正常情况下,可用的电池容量支持负载并为PV提供供电。可用的容量包括2%的关键安全限制,该限制在长期网络故障的情况下保护客户资产。一夜之间,还保持了3%的容量以用于电子电池备份。有关更多信息,请参见https://enphase.com/en-gb/download/iq-battery-5p-usable-capacity-tech-brief。3。生活开始时。4。在低于15°C和大于45°C的温度下,负载功率的降低发生,并且在5°C以下的温度和大于50°C的温度下,放电功率的降低发生降低。5。支持PV分支的所有逆变器,包括Fronius,Solaredge,SMA,Solarmax和Kostal。6。根据地方法规,支持奥地利和德国的阶段失衡管理。7。欧盟(DOC)合格声明的全文可在https://enphase.com/de-de/download/eu-konformitatserklarung上获得。8。无论先到达什么人。限制适用。

IQ电池10TIQ电池10T
1。从交替的电流到电流的交替电流到25°C的标称功率的50%(生命开始时)。实际回报产量可能会根据环境温度,负载模式和其他外部因素而有所不同。2。在日常操作的正常情况下,可用的电池容量支持负载并为PV提供供电。可用的容量包括2%的关键安全限制,该限制在长期网络故障的情况下保护客户资产。一夜之间,还保持了3%的容量以用于电子电池备份。有关更多信息,请参见https://enphase.com/en-gb/download/iq-battery-5p-usable-capacity-tech-brief。3。生活开始时。4。在低于15°C的温度和大于45°C的温度下,负载功率的降低会发生,并且在5°C以下的温度和大于50°C的温度下,放电功率降低。5。支持PV分支的所有逆变器,包括Fronius,Solaredge,SMA,Solarmax和Kostal。6。根据地方法规,支持奥地利和德国的阶段失衡管理。7。欧盟(DOC)合格声明的全文可在https://enphase.com/de-de/download/eu-konformitatserklarung上获得。8。无论先到达什么人。限制适用。
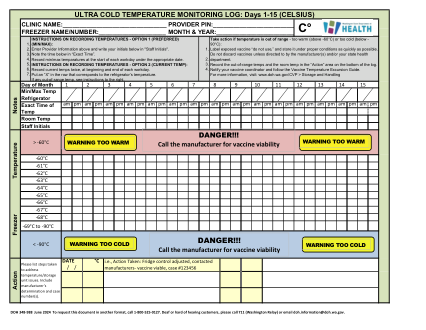
超低温疫苗温度对数-摄氏度
如果温度超出范围,请采取行动 - 温度过高(高于 -60°C)或温度过低(低于 - 90°C):将暴露的疫苗标记为“请勿使用”,并尽快将其存放在适当的条件下。除非制造商和/或您所在州的卫生部门指示,否则不要丢弃疫苗。在日志底部的“操作”区域中记录超出范围的温度和室温。通知您的疫苗协调员并遵循疫苗温度超标指南。有关更多信息,请访问:www.doh.wa.gov/CVP > 存储和处理