XiaoMi-AI文件搜索系统
World File Search System减数分裂
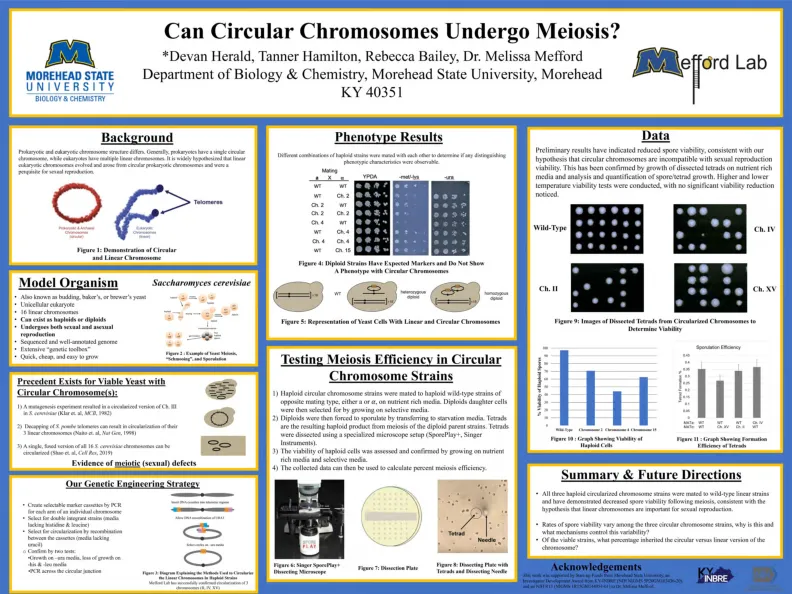
圆形染色体会发生减数分裂吗?
1)单倍体圆形染色体菌株在富含营养培养基上的相反交配类型的单倍体野生型菌株配对。然后,通过在选择性培养基上生长二倍体子细胞。2)然后将二倍体通过转移到饥饿的Me di a来散发。二倍体母体菌株减数分裂是产生的单倍体产物。TETRA DS。3)单倍体细胞的生存能力通过营养素生长

fignl1-firrm对于减数分裂重组至关重要,并防止DNA
control (63) Firrm cKO (649) control (264) Firrm cKO (2937) control (738) Firrm cKO (2777) control (1066) Firrm cKO (1971) control (63) Firrm cKO (649) control (264) Firrm cKO (2934) control (742) Firrm cKO (2811)控制(1082)FIRRM CKO(2015)

1介导减数分裂双链破裂活动
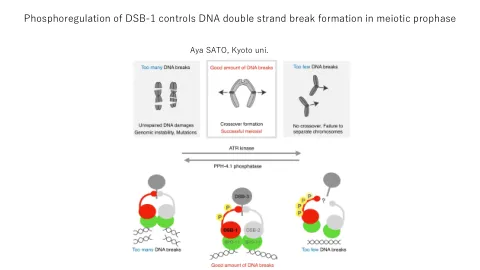
在第一个减数分裂细胞分裂中摘要,大多数生物体的染色体的适当分离取决于chiasmata,这是源自spo11核酸酶催化的编程双链断裂(DSB)的同源染色体之间的连续性交换。由于DSB会导致生殖细胞无法弥补的损害,而缺乏DSB的染色体也缺乏Chiasmata,因此必须仔细调节DSB的数量既不会太高也不太低。在这里,我们表明,在秀丽隐杆线虫中,减数分裂DSB水平受DSB-1的磷酸调节控制,DSB-1是PP4 PPH-4.1磷酸酶和ATR ATL-1 Kinase的相对活性,DSB-1(酵母SPO11辅助剂REC114)的同源物。PPH-4.1突变体中DSB-1磷酸化的增加与DSB形成的减少相关,而DSB-1磷酸化的预防大大增加了PPH-4.1突变体和野生型背景中的减数分裂DSB的数量。秀丽隐杆线虫及其近亲还具有DSB-1的差异旁系同源物,称为DSB-2,而DSB-2的丢失却可以减少年龄增加的卵母细胞中的DSB形成。我们表明,DSB-1的哲学和灭活形式的比例随着年龄的增长和DSB-2的流失而增加,而不可磷酸化的DSB-1则挽救了DSB-2突变体中DSB的年龄依赖性降低。这些结果表明,DSB-2部分进化以补偿DSB-1通过磷酸化的失活,以维持老年动物的DSB水平。我们的工作表明,PP4 PPH-4.1,ATR ATL-1和DSB-2与DSB-1协同作用,以在整个生殖寿命中促进最佳DSB水平。

卵母细胞分化与小鼠的减数分裂程序在遗传上
卵子发生是一种发展计划,通过该计划,配备能力的生殖细胞成为富含施肥的卵。在卵子发生过程中,卵母细胞的生长和分化与减数分裂的起始和进展密切相关。在哺乳动物中,减数分裂起始的时机是性二态性的,只有卵巢且不睾丸生殖细胞在胎儿发育过程中引发减数分裂。因此,胎儿减数分裂开始被认为是随后将卵巢生殖细胞生长和分化为完全生长的卵母细胞的先决条件。在这里,我提供了证据表明,减数分裂的起始和预言I在遗传上与卵母细胞生长和分化是可分开的,因此表明卵子发生在不同的调节下由两个独立的过程组成。这代表了卵子发生程序的新看法,并修改了当前小鼠卵子发生的生殖细胞承诺模型。拟议的修订模型解释了生殖细胞对减数分裂和分化的独立承诺。该模型可以提供有关以前无法解释的女性不育症病例的见解,并对体外卵子发生策略具有实际意义。



利用小鼠天然减数分裂驱动力来抑制入侵种群
入侵啮齿动物是造成环境破坏和生物多样性丧失的主要原因,尤其是在岛屿上。与昆虫不同,包括具有偏向遗传的种群抑制基因驱动在内的遗传生物防治策略尚未在小鼠中开发出来。在这里,我们展示了一种基因驱动策略(t CRISPR),它利用t单倍型的超孟德尔传递来传播单倍型充足的雌性生育基因(Prl)中的失活突变。使用空间明确的基于个体的计算机模拟建模,我们表明t CRISPR可以在一系列现实的基于场的参数值下消灭岛屿种群。我们还设计了转基因t CRISPR小鼠,至关重要的是,它们表现出对修改后的t单倍型和Prl突变的偏向传播,而我们的模型预测这些水平足以消灭它们。这是一个可行的基因驱动系统的例子,用于控制入侵外来啮齿动物种群。

SPO11二聚化控制减数分裂DNA双链断裂的形成
SPO11 二聚化控制减数分裂 DNA 双链断裂形成 Cédric Oger 1 和 Corentin Claeys Bouuaert 1,* 1 鲁汶生物分子科学与技术研究所,鲁汶天主教大学,1348 Louvain-La-Neuve,比利时。 * 通讯地址:corentin.claeys@uclouvain.be。SPO11 通过诱导程序性 DNA 双链断裂 (DSB) 来启动减数分裂重组,但这种催化活性从未在体外重建。在这里,我们使用小小鼠 SPO11 报告了一个重现减数分裂 DSB 形成所有特征的生化系统。我们表明,SPO11 在没有任何伴侣的情况下催化断裂形成,并保持与 5 ¢ 断裂链的共价连接。我们发现 SPO11 的靶位选择受 DNA 底物的序列、可弯曲性和拓扑结构的影响,并提供了 SPO11 可以重新修复单链 DNA 断裂的证据。此外,我们表明 SPO11 在溶液中是单体,而切割需要二聚化才能重建两个混合活性位点。SPO11 及其伴侣 TOP6BL 形成 1:1 复合物,该复合物催化 DNA 切割,其活性与单独的 SPO11 相似。然而,该复合物以更高的亲和力结合 DNA 末端,表明在切割后可能发挥作用。我们提出了一个模型,其中体内 DSB 形成所需的 SPO11 的其他伴侣组装生物分子凝聚物,招募 SPO11-TOP6BL,从而实现二聚化和切割。我们的工作确立了 SPO11 二聚化是控制减数分裂 DSB 诱导的基本机制。

Pristionchus pacificus 减数分裂分析揭示了线虫谱系中同源配对和联会的可塑性
摘要 减数分裂在真核生物中是保守的,但其执行细节各不相同。本文我们描述了一种用于减数分裂分子分析的新型比较模型系统,即线虫 Pristionchus pacificus,它是广泛研究的模型生物秀丽隐杆线虫的远亲。P. pacificus 具有许多解剖学和其他特征,这些特征有助于分析秀丽隐杆线虫的减数分裂。然而,秀丽隐杆线虫失去了减数分裂特异性重组酶 Dmc1 并进化出一种重组独立的机制来使其染色体联会,而 P. pacificus 同时表达 DMC-1 和 RAD-51。我们发现 SPO-11 和 DMC-1 是稳定同源配对、联会和交叉形成所必需的,而 RAD-51 对这些关键的减数分裂过程而言是可有可无的。 RAD-51 和 DMC-1 在减数分裂前期按顺序定位到染色体上,并显示不重叠的功能。我们还展示了 P. pacificus 的新遗传图谱,该图谱揭示了与 C. elegans 非常相似的交叉景观,尽管这些谱系之间在联会和交叉的调节方面存在明显差异。