XiaoMi-AI文件搜索系统
World File Search System刺进


基于 GPS 的飞机进近和着陆系统 (GBAS:地基增强系统) 的开发
尖端技术构筑美好未来:先进宇宙应用技术 隼鸟2号离子发动机及其潜在应用 隼鸟2号——自主导航、制导和控制系统 支持龙宫小行星精确着陆 利用星载激光雷达遥感技术实现隼鸟2号航天器的自主着陆 隼鸟2号:系统设计和运行结果 用于高速、大容量数据通信的卫星间光学通信技术 为三朝深空站开发30kW级X波段固态功率放大器 开发世界最高性能薄膜太阳能电池阵列桨片

通用 DNA 适体可有效抑制 Spike 蛋白/hACE2 相互作用
严重急性呼吸综合征冠状病毒 2 (SARS-CoV-2) 使用其刺突蛋白通过人血管紧张素转换酶 2 (hACE2) 附着到宿主细胞上。可以通过设计一种可以阻断刺突蛋白和 hACE2 之间相互作用的抑制剂来阻止病毒感染。如图 1 所示,一个刺突蛋白三聚体包含三个刺突蛋白,每个刺突蛋白由亚基 S1 和 S2 组成。S1 由 S1A 和 S1B 组成(图 1A),其中 S1B 也称为受体结合域 (RBD),与 hACE2 建立直接相互作用。1 此外,S2 亚基在介导病毒膜与宿主细胞融合方面发挥作用。因此,病毒进入是通过一系列事件完成的,即 S1 与 hACE2 结合,然后触发 S2 将其构象改变为更稳定的融合后状态并允许病毒进入宿主细胞。 2–4 由于 S1 直接与 hACE2 相互作用,许多研究小组一直在积极致力于发现各种生物分子,如抗体 5–10 或适体 11–16,以有效阻断 S1 和 hACE2 之间的相互作用。

高级成像和标记方法破译了脑细胞组织和功能
神经元可以说是生物体中最复杂的形态复杂细胞之一,通常会延伸数百微米(即使不是米)。为了应对其独特的细胞对电池通信能力和空间扩散,神经元已经进化了特定的动态组织和亚细胞分化,例如树突状刺,轴突初始部分或生长锥,以维持特定的功能。在神经系统中,其他细胞类型(例如星形胶质细胞)也表现出特别复杂的形态,适合其特定功能和与神经元共组织。神经元或其他脑细胞的纳米级亚细胞组织以及子隔间之间的元素的运输已成为控制其功能的一组基本特性。突触,尤其是携带大多数兴奋性传播的突触前末端和相关的突触前末端,在突触后受体组织的水平和突触前释放机械1-3的隔离水平上都表现出特别复杂的纳米级组织。通过纳米柱4-7发现,该突触后组织的复杂性增加了。轴突初始段是另一个

超人频道低刺stalk密集的尼奥贝特锂波导通过floquet Engineering
集成的光子芯片逐渐成为信息传输和处理的重要选择,其中集成密度将扮演与综合电路中见证的越来越重要的作用。迄今为止,在制管机上硅晶片已经与低串扰的密集整合做出了巨大的效果,尽管在新兴的二氯甲甲虫在启用锂岩岩(LNOII)平台中仍然非常具有挑战性。在这里,我们报告了一种利用Floquet-Mode-Index调制的策略,以实现宽带零串扰,对LNOI芯片的其他性能指标的影响最小。零串扰的潜在物理学归因于floquet quasienergy的崩溃,这是通过超速频道低cros刺传输的实验性验证的,其多余的损失低。此外,我们在紧凑的LNOI波导阵列中展示了宽带八通道光传输,与传统的波导阵列相比,宽带八通道阵列显示出优势。我们的工作是提高片上光子电路的集成密度的另一种方法,为有希望的LNOI平台中的密集波导应用开辟了不同的可能性。

2020 年末出现多种影响 677 位氨基酸的 SARS-CoV-2 刺突蛋白变体谱系
摘要。严重急性呼吸综合征冠状病毒 2 (SARS-CoV-2) 刺突蛋白 (S) 在宿主细胞进入中起着关键作用。影响 S 的非同义替换并不罕见,并且已在许多 SARS-CoV-2 谱系中固定下来。这些突变的一部分能够逃避中和抗体,或被认为通过增加对细胞进入受体血管紧张素转换酶 2 (ACE2) 的亲和力等机制增强传播。新墨西哥州和路易斯安那州的独立基因组监测计划同时检测到大量 20G 分支(谱系 B.1.2)感染的快速增加,这些感染携带 S 中的 Q677P 替换。该变体于 10 月 23 日首次在美国发现,但在 2020 年 12 月 1 日至 2021 年 1 月 19 日期间,它分别占路易斯安那州和新墨西哥州测序的所有 SARS-CoV-2 基因组的 27.8% 和 11.3%。 Q677P 病例主要在美国中南部和西南部发现;截至 2021 年 2 月 3 日,GISAID 数据显示美国有 499 个该变体的病毒序列。系统发育分析显示至少六个不同的 Q677H 亚谱系独立进化和传播,首次采集日期从 2020 年 8 月中旬到 11 月下旬不等。来自 20G(B.1.2)、20A(B.1.234)和 20B(B.1.1.220 和 B.1.1.222)分支的四个 677H 分支每个分支包含大约 100 个或更少的测序病例,而一对不同的 20G 分支簇分别由 754 个和 298 个病例代表。尽管采样偏差和奠基者效应可能导致了 S:677 多态性变体的出现,但该位置与 S1/S2 边界的多碱基裂解位点的接近性与其在细胞进入过程中的潜在功能相关性一致,表明可能赋予传播或传播优势的特征的平行进化。总之,我们的研究结果表明了同步趋同进化,从而推动了进一步评估 S:677 多态性对蛋白水解加工、细胞趋向性和传递性的影响。

通过免疫信息学设计针对 nCOVID-19 的核衣壳磷蛋白 (N) 和刺突糖蛋白 (S) 的多肽疫苗
核衣壳蛋白 QIGYYRRATRRIRGG HLA-DRB1*11:01 IGYYRRATRRRGGD HLA-DRB1*11:01 GYYRRATRRRIGGDG HLA-DRB1*11:01 TPSTWLTYTGAIKL HLA-DRB1*07:01 DQIGYYRRATRRIRG HLA-DRB1*11:01 PQIAQFAPSASAFFG HLA-DRB1*09:01 WPQIAQFAPSASAFF HLA-DRB1*09:01 QIAQFAPSASAFFGM HLA-DRB1*09:01 IAQFAPSASAFFGMS HLA-DRB1*09:01 AALALLLLDRLNQLE HLA-DRB4*01:01,HLA-DPA1 03:01/DPB1*04, HLA-DRB3*01:0, HLA-DRB1*13:02, HLA-DRB1*11:0, HLA-DRB1*04:04, HLA-DRB1*01:01, HLA-DRB1*04, HLA-DPA1*02:01/DPB1*01:01, HLA-DPA1*01:03/DPB1*02:01, HLA-DRB1*04:05, HLA-DRB1*03:01, HLA-DRB1*08:02, HLA-DRB1*15:01, HLA DQA1*01:01/DQB1*05:01 ALALLLLDRLNQLES HLA-DRB4*01:01, HLA-DPA1*03:01/DPB1*04:02, HLA-DRB3*01:01、HLA-DRB1*13:02、HLA-DRB1*11:01、HLA-DRB1*04:04、HLA-DRB1*04:01、HLA-DRB1*01:01、HLA-DRB1*03:01、HLA-DRB1*04:05、HLA-DPA1*02:01/DPB1*01:01、HLA-DPA1*01:03/DPB1*02:01、HLA-DRB1*08:02、HLA-DRB1*15:01、HLA-DQA1*01:01/DQB1*05:01 PRWYFYYLGTGPEAG HLA-DRB1*07:01 RWYFYYLGTGPEAGL HLA-DRB1*01:01尖峰糖蛋白 AAEIRASANLAATKM HLA-DQA1*05:01/DQB1*03:01 NAQALNTLVKQLSSN HLA-DRB1*11:01 EVFNATRFASVYAWN HLA-DPB1*02:01、HLA DPB1*04:02、HLA-DPB1*05:01、 HLA-DQA1*01:02、HLA-DQA1*05:01、HLA-DQB1*03:01、HLA-DQB1*06:02、HLA-DRB1*01:01、HLA-DRB1*04:04、HLA-DRB1*04:05、HLA-DRB1*07:01、 HLA-DRB1*08:02、HLA-DRB1*09:01、 HLA-DRB1*11:01, HLA-DRB1*15:01, HLA-DPA1*03:01, HLA-DPB1*01:01, HLA-DPA1*01:03, HLA-DPA1*02:01 VFRSSVLHSTQDLFL HLA-DRB1*07:01, HLA-DRB1*01:01, HLA-DRB1*09:01, HLA-DRB1*04:05, HLA-DRB1*04:01, HLA-DRB1*03:01, HLA-DQA1*01:02/DQB1*06:02, HLA-DPA1*03:01/DPB1*04:02, HLA-DRB1*13:02, HLA-DPA1*02:01/DPB1*01:01、HLA-DRB4*01:01、HLA-DQA1*05:01/DQB1*02:01、HLA-DRB1*04:04、HLA- DPA1*01:03/DPB1*02:01、HLA-DQA1*05:01/DQB1*03:01 等位基因 HLA-DRB3*01:01、HLA-DRB4*01:01、HLA-DRB5*01:01 不可用,因此未将其纳入计算。
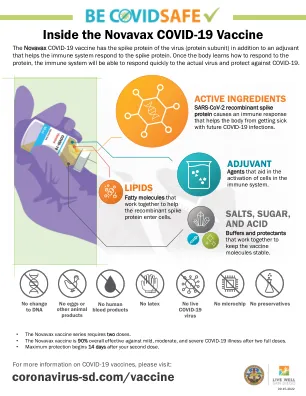
Novavax COVID-19 疫苗内部
Novavax COVID-19 疫苗含有病毒的刺突蛋白(蛋白质亚基),以及帮助免疫系统对刺突蛋白作出反应的佐剂。一旦人体学会如何对蛋白质作出反应,免疫系统将能够快速对实际病毒作出反应并预防 COVID-19。

在流行病期间测量疫苗的有效性
COVID-19 疫情带来的紧迫性促使人们实施了适应特殊情况的临床试验 [ 1 ],并建立了史无前例的公私合作伙伴关系 [ 2 ]。特别是,我们观察到,不同疫苗报告的有效性存在很大差异 [ 3 ],从辉瑞和 Moderna(基于 mRNA)的 * 95% 到阿斯利康的 * 70% 或强生的 * 66%(基于病毒载体)。虽然基于 mRNA 的疫苗(如辉瑞-BioNTech 和 Moderna 疫苗)直接指示我们的细胞产生刺突蛋白,但阿斯利康和强生等病毒载体疫苗采用无害的病毒载体将产生刺突蛋白的遗传指令传递给我们的细胞。一旦产生刺突蛋白,我们的免疫系统就会将它们识别为外来物并产生反应。这包括产生可以结合并中和刺突蛋白的抗体,以及激活 T 细胞。这些免疫反应提供了对病毒的免疫力。鉴于所有这些疫苗都是通过刺激刺突蛋白的产生来诱导免疫反应,值得考虑的是,结果的差异是否可能受到实验条件差异的影响,例如感染者的比例和变异的存在。在本文中,我们展示了一些研究结果,这些研究结果表明,在流行病发展的不同阶段测量疫苗效力可能导致对效力的严重低估。

简介及应用
为Ҷ进一步ՈॆCas13a ⭘于RNA ࠶ᆀ䇺ᯝ的⚥ᓖ,ᕐ䬻઼ -aPes -CROOLQs 䈮仈㓴合ሶ䟽㓴㚊合䞦ᢙ໎ᢰᵟ˄recRPELQase pRO\Perse aPpOLILcaWLRQ,RPA˅઼Cas13a 的旁支活性结合,ᔰ发 ࠪҶާᴹᴤ侩⚥ᓖ的'NARNA ࠶ᆀỰ⍻ᐕާüüS+(R/2C.˄SpecLILc +LJK-SeQsLWLYLW\ (Q]\PaWLc RepRrWer 8Q/2C.LQJ˅。俆ݸ࡙⭘RPA 或R7-RPA ሶṧ૱中的Ṩ䞨࠶ᆀ序列进㹼ᚂᢙ໎,❦ਾ㓿7 䖜ᖅ䞦 䖜ᖅࠪབྷ䟿的RNA ࠶ᆀ,ަ中的目标RNA ࠶ᆀ与crRNA-Cas13 ༽合⢙ 结合◰活Cas13 㳻ⲭ的旁支活性,从而切割ઘത⧟ຳ中࣐的ᣕ࠶ ᆀ,ӗ⭏㜭被Ự⍻的㦗ݹؑਧ。