XiaoMi-AI文件搜索系统
World File Search System前脑

皮质乙酰胆碱动力学可通过胆碱能轴突活动和行为状态预测
人们认为乙酰胆碱 (ACh) 在驱动清醒状态下发生的快速、自发的大脑状态转变方面发挥着作用;然而,这些状态变化期间皮质 ACh 活动的时空特性仍不清楚。为了解决这个问题,我们同时对 GRAB-ACh 传感器、表达 GCaMP 的基底前脑轴突和行为进行成像。我们观察到在运动和瞳孔扩张期间轴突和 GRAB-ACh 活动之间存在高度相关性。仅从轴突活动就可以准确预测 GRAB-ACh 荧光,并且局部 ACh 活动在距离轴突较远的地方会降低。对 GRAB-ACh 轨迹进行反卷积使我们能够解释传感器动力学并强调快速清除小 ACh 瞬变。我们训练了一个模型来根据瞳孔大小和跑步速度预测 ACh,

走向生命机器:触觉感知、抓握和社交机器人的现状和未来趋势
1 技术创新研究所 (TII),阿布扎比,阿拉伯联合酋长国 2 比萨大学“E. Piaggio”研究中心,意大利比萨 3 利兹大学,计算机学院,利兹 LS2 9JT,英国 4 布里斯托大学工程学院工程数学系和布里斯托机器人实验室,布里斯托,英国 5 巴斯大学工程与设计学院电子电气工程系,巴斯,英国 6 本研究部分由英国工程与物理科学研究委员会根据 EP/V052659/1 号资助。 7 本研究部分由利华休姆研究领导奖“用于机器人触觉的仿生前脑”(RL-2016-39) 资助。 8 作者已确认,本研究中所有可识别的参与者均已同意发表。∗ 任何通讯均应寄给作者。

抑制素-2 在疾病中的作用
抑制素 (PHB) 是一类保守的蛋白质,主要位于线粒体内膜和细胞核中,具有多种生物学功能。PHB 包括 PHB1 (32 kDa) 和 PHB2 (37 kDa),是包含口素和浮素的超家族成员 [1]。PHB 主要以具有玫瑰花结结构的聚合物形式存在,直径为 20–25 nm [2,3]。据推测,PHB 调节细胞周期、衰老和肿瘤抑制,同时特异性地抑制 DNA 合成的起始 [4]。PHB1 抑制几种类固醇激素受体的信号传导 [5-7],而 PHB2 选择性地抑制雌激素受体 (ER) 活性 [8]。 PHB2 参与重要的细胞过程,包括细胞存活、凋亡、代谢、炎症、基因转录和信号转导 [9,10]。已发现秀丽隐杆线虫和小鼠的 PHB 缺失会导致胚胎死亡 [11,12]。为了进一步阐明 PHB2 在生理和病理生理过程中的作用,已经建立了几种组织特异性 PHB2 敲除小鼠模型。例如,前脑特异性 PHB2 缺陷小鼠表现出 tau 过度磷酸化和神经退行性[13],肾足细胞中 PHB2 的缺失导致肾小球硬化[14],肝细胞特异性 PHB2 敲除小鼠表现出肝功能衰竭和糖异生受损[15],β 细胞特异性 PHB2 敲除导致 β 细胞功能受损和糖尿病[16]。心脏特异性 PHB2 敲除小鼠会出现心力衰竭并死亡[8]。结果表明正常器官功能需要 PHB2。PHB1 或 PHB2 中的任何一个的敲低都会导致另一个的敲低,从而造成 PHB“超复合物”表达降低。例如,在前脑或 HEK293T 细胞中,其组装伙伴 PHB1 的有效丧失伴随 PHB2 的耗竭 [ 13 , 14 ],这突显了 PHB 亚基在某些组织中的功能相互依赖性。

exelon®解决方案
痴呆症中的病理变化,例如阿尔茨海默氏病,涉及从基础前脑到大脑皮层和海马的胆碱能神经元途径。这些途径已知与注意力,学习和记忆以及其他认知过程有关。ivastigmine是一种脑选择性的乙酰乙酰基 - 乙酰胆碱酯酶抑制剂,被认为可以通过功能完整的胆碱能神经元释放的乙酰胆碱的降解来促进胆碱能神经传递。来自动物研究的数据表明,利瓦斯汀明选择地增加了皮质和海马中乙酰胆碱的可用性。因此,Exelon可能对与阿尔茨海默氏病和帕金森氏病有关的胆碱能介导的认知缺陷有改善作用。此外,还有一些证据表明,胆碱酯酶抑制作用可以减慢淀粉样蛋白生成β-淀粉样蛋白蛋白(APP)片段的形成,因此淀粉样蛋白斑块是阿尔茨海默氏病的主要病理特征之一。

我们能解释数千种分子鉴定的小鼠神经元类型吗?从知道到理解
脑计划细胞普查网络 (BICCN) 于 2023 年 12 月 13 日在《自然》杂志上发布了《全鼠脑图谱》出版包(https://www.nature.com/collections/fgihbeccbd,2024 年 5 月 5 日访问)。这项单细胞转录组、表观基因组和空间转录组综合工作将小鼠脑中存在的不同神经元细胞类型的数量更新为惊人的总数,略多于 5300 种,揭示了它们的分子多样性与它们的相对位置一致。我们在此提出的问题是:我们能否解释如此多不同类型的细胞是如何产生和定位的?这个问题与另一个问题相关:我们是否有形态模型允许在相对位置和神经元类型规范方面将这种程度的多样性相关联?令人惊讶的是,答案是可能的,而且几乎是肯定的。 BICCN 出版物隐含地使用了 Herrick 的传统柱状脑模型([ 1 ];图 1 a-d),可能是 Swanson 的修改版([ 2 , 3 ];图 1 e),或 Dong [ 4 ] 在 Allen 研究所的成年小鼠大脑图谱 [mouse.brain-map.org] 中使用的模型。该模型将端脑、间脑、中脑、后脑和脊髓视为主要分区(五个喙尾小泡;图 1 a)。在该模型中,Herrick 的最小单位由四个功能实体表示(脑干和脊髓中定义的躯体运动、内脏运动、内脏感觉和躯体感觉柱:Sm、Vm、Vs、Ss;图 1 a、d)。本文作者将它们外推到前脑(即间脑的 Eth、Dth、Vth、Hth;端脑的 Hi、Pir、Str、Se;图 1 a-c 中统一颜色的代码)。请注意,前脑柱可能执行与后脑不同的功能,尽管间脑在功能上被解释为脑干的延续。总的来说,这就构成了 5 个囊泡 × 4 个柱 = 20 个柱状单元,它们应该产生最近发现的 5300 种神经元类型(平均每柱 265 种细胞类型)。

KCNQ1的内含子1包含一个调节区域,该区域...
图1。识别KCNQ1基因座中预测的调节元件。a)UCSC基因组浏览器视图描述了第一个内含子使用情况不同的KCNQ1的两个同工型,而KCNQ1OT1则是该位点中长的非编码RNA。同工型下方的轨道表示来自GWA的QT间隔相关的SNP,其位置在所有轨道中都延伸到灰色条上。出现的轨道描绘了基于心脏特异性数据集的预测调节元件,最低的三个轨道描绘了组蛋白公开可用的芯片seq实验的测序读取,标志着来自两个人类左心室的H3K27AC,并留下ATRIA ATRIA ATAC-SEQ-SEQ-seq实验。b)小鼠中KCNQ1基因座的UCSC基因组浏览器视图,其先前表征的远景增强子在KCNQ1的内含子1中。较低的两个曲目描绘了从胚胎第15天公开可用的小鼠心脏和前脑的ATAC-SEQ数据集的测序读数。
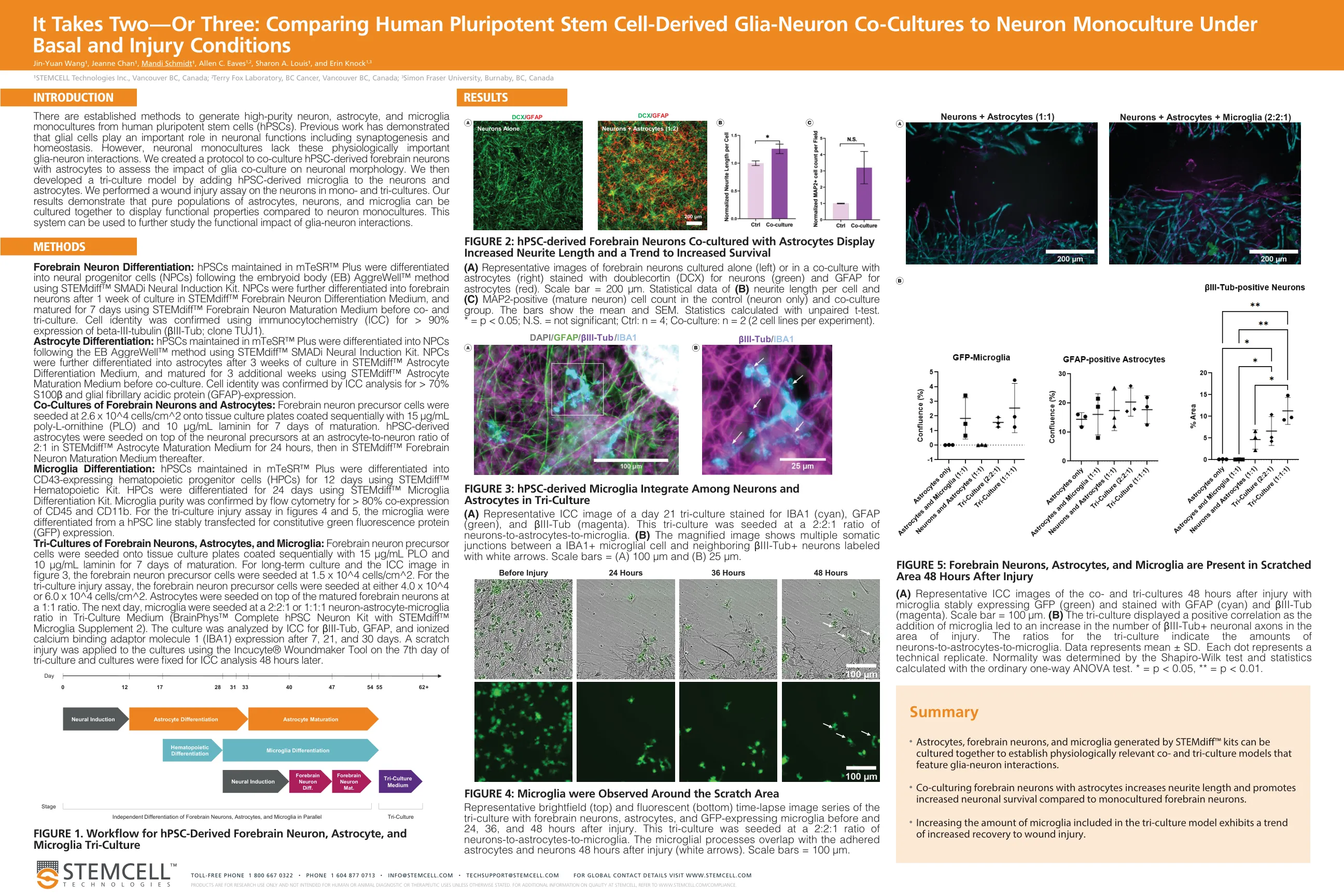
比较人类多能干细胞得出的胶质细胞 -
有建立的方法来产生人类多能干细胞(HPSC)的高纯性神经元,星形胶质细胞和小胶质细胞。先前的工作表明,神经胶质细胞在神经元功能中起重要作用,包括突触发生和稳态。然而,神经元单栽培缺乏这些在生理上重要的神经神经元相互作用。我们创建了一种与星形胶质细胞共同培养HPSC衍生的前脑神经元共同培养的方案,以评估神经胶质共培养对神经元形态的影响。然后,我们通过将HPSC衍生的小胶质细胞添加到神经元和星形胶质细胞中,开发了三个文化模型。我们对单培养的神经元进行了伤口损伤测定法。我们的结果表明,与神经元单栽培相比,可以一起培养星形胶质细胞,神经元和小胶质细胞的纯种群以显示功能特性。该系统可用于进一步研究胶质神经元相互作用的功能影响。

全脑脑
全脑脑是复杂的大脑畸形,这是由于早期胎儿发育过程中大脑不完全的裂解而导致的。这种情况的特征在于普罗德龙(胚胎的前脑)的失败,以正确分成大脑半球的双叶,导致影响大脑和面部特征的异常。根据大脑分裂的严重程度,全脑脑分为四种类型:Alobar Holoporsencephaly:最严重的形式,其中没有脑半球分离,导致单个脑室心室和一个单裂脑。半月骨全脑脑:大脑半球部分分离,大脑的结构在某种程度上介于Alobar和Lobar之间。Lobar Holoporsencephaly:最少的严重形式,具有更好的脑半球分离和更正常的大脑结构。中半球间变体(syntelcephaly):半球在大脑中间没有分离,但可能在前和后方面更正常地分裂。是什么导致全脑脑?

使用开放数据的胎儿功能MRI自动脑掩盖
抽象的胎儿静止状态功能磁共振成像(RS-FMRI)已成为表征出生前脑发育的关键新方法。尽管这种方法的快速而广泛地增长,但目前,我们缺乏适合解决该数据类型固有的独特挑战的神经影像处理管道。在这里,我们解决了最具挑战性的处理步骤,在数千个非平稳的3D大脑体积中,胎儿大脑与周围组织的快速而准确地隔离。利用我们的1,241个手动追踪的胎儿fMRI图像的图书馆,我们培训了一个卷积神经网络(CNN),该卷积神经网络(CNN)在来自单独的扫描仪和人群的两个持有测试集中实现了出色的性能。此外,我们将自动屏蔽模型与现有软件的其他fMRI预处理步骤结合在一起,并洞悉我们对每个步骤的改编。这项工作代表了针对全面的,开源的工作流程的最初进步,并具有公开共享的代码和数据,用于胎儿功能性MRI数据预处理。

OTX和OTP基因在大脑发育中的作用
摘要在过去十年中,通过一系列动物模型在功能水平上鉴定并表征了许多参与大脑诱导,规范和区域化的基因。在这些基因中,OTX1和OTX2,果蝇正畸形(OTD)基因的两个鼠类同源物,编码转录因子,在鼻脑的形态发生中起关键作用。经典的敲除研究表明,OTX2对于早期规范和随后的前神经板的维护至关重要,而OTX1主要是正常的皮质生成和感官器官发育所必需的。将两个基因产物的最小阈值正确地构成前脑的构图和地质组织者的定位。第三基因,骨科(OTP)是控制神经内分泌下丘脑发育的遗传途径的关键要素。本综述介绍了OTX1,OTX2和OTP函数的全面分析,以及otx基因被果蝇同源物otd的模型所暗示的可能的进化含义。