XiaoMi-AI文件搜索系统
World File Search System双链


双纳米
“当第一个人拿棍子打倒香蕉时,第二个人立刻就想出了如何借助这根棍子把香蕉拿走。所有真正的新技术都是双重用途的,”Rusnano 董事会主席顾问 Vasily Grudev 开玩笑说。对于高科技公司来说,军方是理想的客户。他们拥有稳定的、通常是大量的资金;他们比平民消费者更有可能需要最好的,而不是最便宜的。从这个意义上说,俄罗斯国防部仅2013年就公布了2.1万亿美元的预算。rub.,令人愉快的“也不例外。”有趣的是,世界各地的军事界对纳米技术的兴趣与日俱增。冷战后的太空竞赛化为泡影,许多有前途的武器因其破坏力而被禁止,而所谓常规武器的总体轮廓自第二次世界大战以来并没有发生根本性的变化——所有这些同样的坦克、飞机、舰艇……就连美国在“常规”武器领域也走上了现代化改进的道路,不断增加战斗力并提高现有装备的战术和技术特性。简而言之,军备竞赛已转向拯救士兵的生命。要做到这一点,就必须让他在战场上停留的时间更短,更安全。因此,无人驾驶车辆、工兵机器人、侦察机器人、减轻和强化装甲的项目蓬勃发展……换句话说,电子和材料科学是纳米技术影响尤其巨大的行业。例如,很明显,寻找新合金不会带来复合材料实验或晶格水平变化所承诺的突破性结果。如今,陶瓷装甲已成功与金属装甲展开竞争。这个市场上有前途的俄罗斯企业是来自新西伯利亚的 NE-VZ-Ceramics 公司。它由 Rusnano 和 NEVZ-Soyuz 控股公司于 2011 年创建。生产装甲陶瓷,用于防弹衣和装备防护。产品已通过俄罗斯及国外测试。其明显的优点是重量轻、防护性能高,但也有“侧面”的优点。陶瓷更难被雷达探测到,并且不太容易被寻的弹药探测到。这为其在


双糖尿病
本评论探讨了双糖尿病的病理生理学,临床意义和管理。肥胖,久坐的生活方式和遗传易感性的越来越多的患病率模糊了1型和2型糖尿病之间的差异,从而导致诊断性和治疗性挑战。双糖尿病均表现出两种糖尿病类型的重叠症状,因此准确的诊断至关重要。生物标志物,例如C肽水平,自身抗体测试和胰岛素抵抗标记,有助于将双糖尿病与经典糖尿病亚型区分开。早期干预是必要的,因为这种病的微血管和大血管后果的风险升高,例如视网膜病变,肾病和心血管疾病。有效管理整合了药理学和生活方式的方法。二甲双胍,葡萄糖共转运蛋白2(SGLT2)抑制剂,胰高血糖素样肽-1(GLP-1)受体激动剂和胰岛素治疗调整所有促进血糖控制和代谢结果。此外,结构化运动,饮食修饰和体重管理对于降低胰岛素抵抗和保留β细胞活性至关重要。精密医学,人工智能(AI)驱动的医疗保健和连续葡萄糖监测(CGM)的潜力为个性化治疗策略提供了有希望的进步。未来的研究应集中于有针对性的免疫疗法,基因分析和精致的临床指南,以改善早期检测和个性化治疗,并具有长期结局。审查强调需要采用多学科方法来管理双重糖尿病,确保早期诊断,优化治疗和改善代谢健康以减轻长期并发症。

使用双...
伊朗Semnan University电气和计算机工程学院电气工程系。b卫生信息管理和技术系,伊朗喀山喀山医学科学学院,伊朗喀山。C喀山医学大学伊朗喀山喀山医学大学的健康信息管理研究中心。D研究所Digihealth,Neu-ULM应用科学大学,德国Neu-Ulm。orcid ID:A。M. Nickfarjam https://orcid.org/0000-0000-0003-3782-3038摘要。我们提出了用于分割和分类脑肿瘤的U-NET体系结构的修改版本,从而引入了向下采样和向上采样之间的另一个输出。我们建议的体系结构利用了两个输出,在分割输出旁边添加了分类输出。中心想法是在应用U-NET的上采样操作之前使用完全连接的图层对每个图像进行分类。这是通过利用在下采样过程中提取的功能并将其与完全连接的层相结合的分类来实现的。之后,通过U-NET的上采样过程生成分段图像。初始测试对骰子系数,准确性和敏感性分别为80.83%,99.34%和77.39%的可比模型显示了竞争性结果。这些测试是在2005年至2010年的中国广州Nanfang医院,中国广州Nanfang医院和中国天津医科大学的综合医院的数据集上进行的,其中包含3064个脑肿瘤的MRI图像。

思科维持对立与美国 - 戴链链 - ...
思科在各个方面(包括供应链网络安全)中仍然是网络安全先驱和行业合作伙伴。我们的价值链安全和供应链计划擅长采用和部署最佳实践,并将继续与我们的客户合作,以确保满足这些标准。Cisco将继续保持一致,并促进与网络安全的信任和透明度提高,以更好地保护我们的业务和客户有关的行业标准。

SCOR参考模型作为建筑链链管理的工具
本文介绍了由供应链委员会提出的SCOR参考模型的基本结构,该模型被广泛用作协助供应链管理的工具,并讨论了其在建筑链管理中实施的机遇和困难。为此,已经详细阐述了有关GCS文献以及与建筑有关的主要挑战的简要回顾。接下来,从概念基础来看,对一家公司的半结构化访谈进行了陶瓷块问题,到结构性砖石公寓企业,目的是使在混凝土情况下与SCOR建模时可能出现的问题相关联。对新供应链建模技术的研究可以帮助许多不知道其供应问题的公司以寻求解决方案。

利用连续 CRISPR/Cas9 基因编辑技术对神经元蛋白进行双链标记和操作
CRISPR/Cas9 介导的基因敲入方法能够标记单个内源性蛋白质,从而如实地确定它们在细胞中的时空分布。然而,由于编辑事件之间存在串扰,因此在神经元中可靠地多路复用基因敲入事件仍然具有挑战性。为了克服这个问题,我们开发了条件性激活基因敲入表达 (CAKE),从而实现高效、灵活和准确的多路复用基因组编辑。为了减少串扰,CAKE 基于顺序重组酶驱动的向导 RNA (gRNA) 表达来控制每个供体序列的基因组整合时间。我们表明,CAKE 广泛应用于大鼠神经元,以共标记各种内源性蛋白质,包括细胞骨架蛋白、突触支架、离子通道和神经递质受体亚基。为了充分利用 CAKE,我们使用超分辨率显微镜解决了内源性突触蛋白的纳米级共分布,表明它们的共组织与突触大小相关。最后,我们引入了可诱导二聚化模块,可精确控制活神经元中的突触受体动力学。这些实验凸显了 CAKE 揭示新生物学见解的潜力。总而言之,CAKE 是一种多功能的多重蛋白质标记方法,可以检测、定位和操纵神经元中的内源性蛋白质。

小分子靶向疗法诱导残留肿瘤细胞中对DNA双链破裂修复的依赖性
用靶向疗法生存的残留癌细胞充当最终抗性疾病的储层。尽管对靶向残留细胞的治疗非常感兴趣,但由于我们对这种细胞状态中存在的脆弱性的有限了解,努力受到阻碍。Here, we report that diverse oncogene-targeted therapies, including inhibitors of epidermal growth factor receptor (EGFR), anaplastic lymphoma kinase (ALK), KRAS, and BRAF, induce DNA double-strand breaks and, consequently, ataxia-telangiectasia mutated (ATM)–dependent DNA repair in oncogene-matched residual tumor cells.在细胞系,小鼠异种移植模型和人类患者中观察到的这种DNA损伤反应是由涉及胱天蛋白酶3和7激活的途径以及下游caspase激活的脱氧核糖核酸酶(CAD)的驱动的。CAD又通过其内源性抑制剂ICAD的caspase介导的降解而激活。在EGFR突变非小细胞肺癌(NSCLC)的模型中,通过小分子EGFR靶向治疗的肿瘤细胞合成依赖于ATM,并与ATM激酶抑制剂在体内消除这些细胞。这导致EGFR突变体NSCLC小鼠异种移植模型的渗透性和耐用反应更多,包括源自已建立的细胞系和患者肿瘤的响应。最后,我们发现,具有携带共同发生的EGFR突变体NSCLC的罕见患者,ATM中的功能丧失突变在第一代EGFR抑制剂疗法中与EGFR突变NSCLC患者缺乏缺乏有害ATM突变的患者相对于第一代EGFR抑制剂疗法表现出扩展的无进展生存率。一起,这些发现为基于机制的ATM抑制剂与现有靶向疗法的基于机制的整合建立了理由。
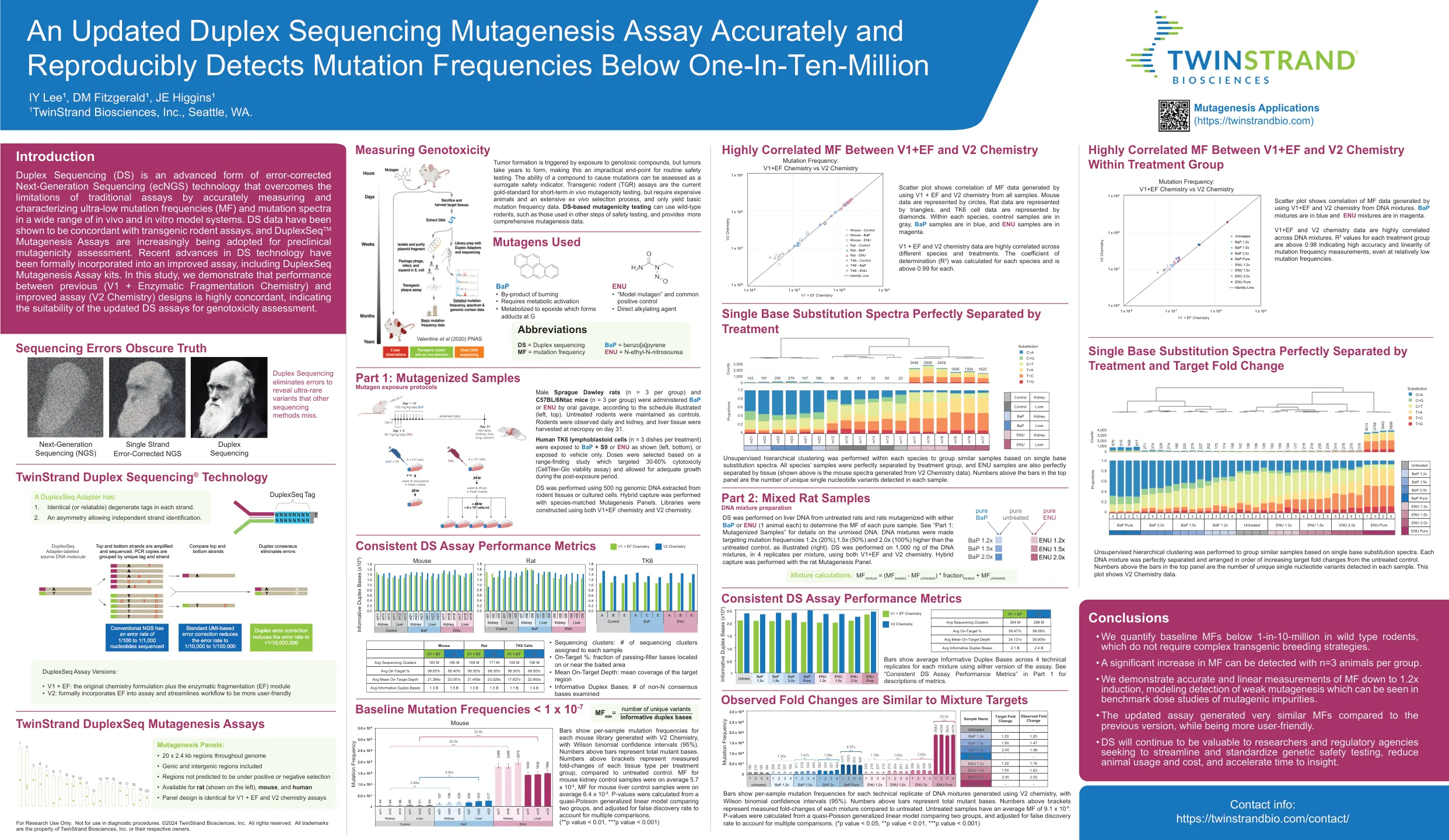
准确的更新的双链测序诱变测定法,可重复检测到低于一十分三百万的突变频率
条显示了用V2化学产生的每个小鼠文库的每样本突变频率,威尔逊二项式置信区间(95%)。条上方的数字代表总突变碱基。与未处理的对照相比,支架上方上方的数字代表每个治疗组的每种组织类型的倍数变化。MF平均为5.7 x 10 -8,小鼠肝对对照样品的MF平均为6.4 x 10 -8。p值是从比较两组的准散孔概括的线性模型中计算得出的,并根据错误的发现率进行了调整以考虑多个比较。(** p值<0.01,*** p值<0.001)仅用于研究使用。不适用于诊断程序。©2024 Twinstrand Biosciences,Inc。保留所有权利。所有商标都是Twinstrand Biosciences,Inc。或其各自所有者的财产。