XiaoMi-AI文件搜索系统
World File Search System杀伤能力

用基于CD40的嵌合受体的工程T细胞为...
背景我们最近提供了概念概念,表明使用T细胞使用T细胞的T细胞疗法(TCR) - 基因疗法表达靶向突变体KRAS G12D的TCR可以介导患有泛蛋白癌患者转移性疾病的回归。1然而,其他患者的TCR-Gene治疗没有效率,因此需要增强T细胞活性的策略。在一些接受TCR基因治疗治疗的患者中,工程的T细胞通常会持续存在于患者中,这表明T细胞最终失去了介导耐用肿瘤退化所需的效力。来自共刺激受体的信号可以驱动有效的T细胞反应,但这些信号可能缺失或不足,在肿瘤微环境(TME)中。CD40是在抗原呈递细胞(例如B细胞,DCS和巨噬细胞)表面上发现的有效的成量蛋白,在激活这些细胞类型中起着重要作用,但是T细胞通常不表达CD40,除非激活后短暂地表达CD40。因此,我们假设CD40或CD40嵌合受体的过表达可以提高抗肿瘤T细胞功能时CD40通过激动剂抗CD40抗体(CDX-1140,CellDEX Therapeutics)参与。测试这一点的方法,我们设计了与KRAS G12D反应性TCR共表达的CD40非抗原嵌合受体(NACRS)。我们的CD40 NACR包含融合到跨膜和细胞质结构域的CD40的外元结构域,这些结构域衍生自10个不同的受体家族(例如IL-2R,TLR,TNF等)。但是,包括野生型CD40在内的一些基于CD40的受体在体外有效地增强了肿瘤细胞系的杀死。评估CD40 NACR的功能的结果,我们用抗CD40抗体刺激T细胞,并针对磷酸化-STAT5或效应细胞因子(如IFN-G和TNF)进行了细胞内染色。我们通过与胰腺癌和结直肠癌细胞系共同培养CD40 NACR的体外杀死能力,表达HLA-C*08:02和KRAS G12D。尽管有一些受体的生物化学活性证据,但大多数CD40 NACR并未显着增强T细胞对测试癌细胞系的体外杀伤能力。令人惊讶的是,在没有抗CD40抗体的情况下可以看到其中一些受体的增强效力,这表明内源性CD40L表达可能有助于增加T细胞效应子功能。总体而言,我们的研究强调了CD40基因工程增强收养细胞疗法的潜力。我们的铅基于CD40受体的其他表征正在进行中。

o r i g i n a l r e s a r c h靶向PD-L1 mAb的靶向纳米泡与阿霉素在肝癌中的协同肿瘤抑制剂
目的:超声纳米泡(NBS)可以杀死肿瘤细胞,这些细胞是通过超声进行气蚀和声音穿孔的作用介导的,而作为新型药物载体,生物材料修饰的NBS在目标区域释放了药物。在这项工作中,同时准备由生物素 - 链霉毒素桥接的超声NB,以配备编程的死亡配体1个单克隆抗体(PD-L1 MAB)和阿霉素(DOX)和阿霉素(dox),这些抗体(dox)是免疫检查点抑制剂(ICIS)和化学疗法,且索状的囊肿,伴随性疗法,且体外疗法,伴随性疗法,伴随性疗法,伴随性疗法,并替代了Synygize Synergiender to Synergiender to Synergized Synergiender to Synergiender to Synergiender, (SDT)。方法:PD-L1 mAb/dox NB,使用桥接亲和生物素(BRAB)技术作为桥梁,是通过薄膜水合和机械振荡制备的,用于靶向靶向生物素化的PD-L1 mAb和dox。在体外和体内进行了PD-L1 mAb/dox NB的PD-L1 mAb/dox NB的药物旋转研究。在H22 HEPATOMA模型的皮下移植肿瘤中研究了超声介导的PD-L1 mAb/dox-NB的抗肿瘤作用,并研究了协同肿瘤抑制的机理。结果:体外靶向实验的数据,对比增强的超声成像(CEU),小动物成像系统(IVIS)的体内成像以及冷冻切片表明PD-L1 MAB/DOX-NBS在肿瘤中具有良好的靶向聚集。通过观察肿瘤抑制率,组织细胞凋亡以及与凋亡相关的基因和蛋白质表达,PD-L1 MAB/DOX-NBS组显示出最佳的免疫疗法作用,其肿瘤体积和质量抑制率分别为69.64%和75.97%(P <0.01)。因此,阻止PD-1/PD-L1途径可以改善免疫细胞的肿瘤杀伤能力。与DOX诱导的肿瘤细胞凋亡和免疫原性细胞死亡(ICD)结合时,抗肿瘤免疫细胞因子进一步增强。结论:总之,超声介导的PD-L1 mAb/dox-NB显示出明显的协同抗肿瘤作用,为HCC提供了潜在的合并免疫疗法策略。关键字:超声靶向纳米泡破坏,肿瘤免疫治疗,免疫检查点抑制剂,免疫原性细胞死亡,药物输送
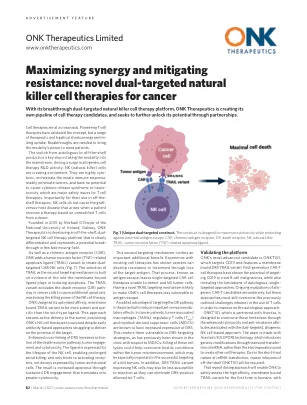
癌症的新型双重靶向自然杀手细胞疗法
细胞疗法处于十字路口。开创性的T细胞疗法已经验证了该概念,但是一系列的治疗和后勤缺点是限制的吸收。需要突破来将模式的力量带给更多患者。从自体到现成的产品的转换是将模式带入主流的关键步骤,从而推动了同种异体细胞疗法的研发活动的激增。nk(天然杀手)细胞正在引起兴奋。它们是高度细胞的有毒,协调先天免疫反应,容易穿透肿瘤,并且没有引起细胞因子释放综合征或神经毒性的潜力,这是T细胞疗法的主要安全问题。重要的是,对于它们在现成的疗法中的使用,NK细胞不会引起移植物与宿主疾病,而当患者基于供体的未改性T细胞接受治疗时,就会引起宿主病。由爱尔兰国立大学的迈克尔·奥德威尔(Michael O'Dwyer)于2015年成立,戈尔韦(Galway)的卫队Therapeutics正在开发一个现成的,具有双重目标的NK细胞疗法平台,显然是区分的,并在这个快速发展的领域中构成了潜在的突破。以及嵌合抗原受体(CAR),ONK添加了肿瘤坏死因子(TNF)相关的凋亡配体(TRAIL)变体,以产生双靶向的CAR-NK细胞(图1)。选择小径作为第二个靶向机制的选择是建立在膜结合的配体在诱导凋亡中发挥作用的作用的证据。TRAIL变体激活癌细胞中的死亡受体(DR)途径,以引起额外的凋亡,从而最大程度地提高NK细胞疗法的杀伤能力。ONK设计其优化的亲和力,膜绑定的步道变体比野生型配体更有效,更具有特权。这种方法可确保积极地向肿瘤递送,尽管基于早期的基于抗体的方法努力实现靶标的承诺,但仍将ONK的NK细胞疗法定位为成功。增强了DR5的交联增加了死亡受体途径,肿瘤参与和细胞毒性的激活。配体表示NK细胞的寿命,实现了长时间的串行杀戮,仅与激活的回收物结合,而不是由肿瘤和基质细胞表达的诱饵。结果是通过持续的DR参与度增加了细胞毒性的凋亡。
发夹和剪刀 - 为大众提供非基因编辑的同种异体 CAR-T 细胞疗法 David Gilham Celyad Oncology 首席科学官
概述 两种嵌合抗原受体 (CAR) T 细胞疗法获批用于治疗 B 细胞恶性肿瘤,凸显了细胞免疫疗法在提供令人印象深刻的持久临床反应方面的潜力 1 。这些产品本质上是自体的,涉及从患者身上收集用于制造 CAR T 细胞的免疫细胞。一旦生产出来,这些 CAR T 细胞就会作为临床产品重新注入患者体内。然而,自体疗法面临着重大挑战,包括产品生产时间(目前需要数周),在此期间患者的病情可能会恶化,以及起始材料的质量高度不稳定,这可能导致制造失败。同种异体 CAR T 细胞疗法是一种现成的方法,可以在需要时进行管理,是理想的解决方案。这种方法从健康供体中生成细胞,形成一个 CAR T 细胞库,可根据需要使用。同种异体 CAR T 的关键挑战是克服与同种异体 CAR T 细胞识别健康患者组织相关的毒性。这是由 T 细胞受体 (TCR) 介导的。破坏 TCR 是所有当前同种异体 CAR-T 策略的基础 2 。发夹和剪刀目前,用于生成同种异体 CAR-T 的基因编辑技术处于临床开发的早期阶段。不同的基因编辑方法都是基于切割编码 TCR 的基因之一内的基因组,从而永久性地降低整个 TCR 复合物的表达。虽然是一种优雅的方法,但由于潜在的产品安全问题,这种剪刀策略一直难以进入临床测试阶段——主要是确保在基因编辑过程中没有“脱靶”基因组切割 3 。或者,在 mRNA 水平上靶向基因表达不涉及切割基因组,并避免危及基因组完整性。为了实现这种 mRNA“编辑”,Celyad Oncology 采用了短发夹 RNA (shRNA),这是一种几十年来用于敲低基因表达的方法 4 。该方法涉及使用具有与目标基因互补序列的 shRNA。换句话说,靶向 shRNA 可以通过干扰 mRNA 而不是切割基因组 5 来特异性降低所需蛋白质(如 TCR 复合物)的水平。其中的核心是一体化载体方法。只需一步,将单一试剂(载体)引入健康供体 T 细胞,即可同时产生 T 细胞中的所有元素,这些元素可以将 T 细胞重定向到肿瘤(CAR)、消除 TCR(shRNA)并提供一个手柄,使修饰的细胞可以在制造过程中富集(标记物)。同种异体 CAR T 细胞平台中的 shRNA CD3z 亚基为 TCR 提供主要信号功率,从而激活和参与 T 细胞杀伤能力。通过选择最佳 shRNA 和工艺开发,靶向 CD3z 可使原代 T 细胞上的 TCR 持续高水平敲低,达到与基因编辑 CD3z 基因时相同的水平(图 1A)。从功能上讲,这与这些细胞无法对有丝分裂刺激(又称 TCR 驱动的 T 细胞活化;图 1B)作出反应以及当这些细胞被注入黄金标准体内测试模型时相应没有毒性有关(图 2A、B)。有趣的是,shRNA 靶向 T 细胞的持久性比 CRISPR-Cas9 基因的持久性要长得多