XiaoMi-AI文件搜索系统
World File Search System杀虫

恶性疟原虫组氨酸富集蛋白 2 缺失的遗传特征及其对印度奥里萨邦疟疾干预的影响
摘要。通过恶性疟原虫 (P. falciparum) 富含组氨酸的蛋白质 2 (pfhrp2) 基因缺失而导致的诊断逃逸是全球消除疟疾工作的主要潜在障碍。我们调查了印度奥里萨邦 15 个疟疾流行村 pfhrp2 基因缺失的流行情况,并模拟了它们对正在进行的国内疟疾干预计划的影响。我们发现 61.6% 的亚潜伏性恶性疟原虫感染(即快速诊断测试 [RDT] 阴性和聚合酶链反应 [PCR] 阳性)有 pfhrp2 基因缺失,这些缺失主要位于外显子 2 区域(96.2%),并且主要在发热个体的样本中发现(82.6%)。在携带完整 pfhrp2 外显子 2 基因座的亚专利感染个体样本子集中,我们对 DNA 测序和蛋白质多样性特征进行了表征。我们的分析揭示了新的氨基酸重复基序(231 – 293 个氨基酸),这些变异重复序列与 RDT 1 /PCR 1 样本的重复序列不同。我们还在 pfhrp2 基因缺失的背景下评估了国家资助的大规模筛查和治疗干预。我们发现,与单独进行 RDT 治疗相比,大规模筛查和治疗结合其他干预措施(例如分发长效杀虫蚊帐、室内滞留喷洒)降低了携带 pfhrp2 缺失的恶性疟原虫(调整后的相对风险比 [aRRR] = 0.3;95% CI = 0.1 – 1.0)和携带完整 pfhrp2 基因的恶性疟原虫(aRRR = 0.4;95% CI = 0.2 – 1.1)的相对感染风险。总之,我们的研究结果强调,在印度朝着 2030 年消除疟疾的目标迈进之际,需要替代的诊断目标和工具。

植物相关微生物在植物健康和发育中的作用:以沙雷氏菌属为例
摘要 沙雷氏菌属是肠杆菌科的一种菌种,存在于多种生态环境中。近年来,沙雷氏菌已成为促进植物生长和防御植物病虫害的多方面贡献者。本综述探讨了沙雷氏菌诱导植物生长和缓解非生物和生物胁迫的机制。沙雷氏菌与植物生态系统的无缝整合使沙雷氏菌能够产生群体感应分子 N-酰基高丝氨酸内酯 (AHL),促进植物组织的定植并利用植物分泌物中的营养。这种错综复杂的通讯网络使沙雷氏菌能够产生植物激素并分解土壤中的必需营养物质供植物吸收。面对生态竞争对手,许多沙雷氏菌菌株表现出非凡的适应性,产生多种水解酶和抗菌、抗真菌或杀虫化合物,有效控制有害细菌、真菌和害虫。此外,有益的沙雷氏菌菌株还分别使用诱导系统抗性 (ISR) 和耐受性 (IST) 来缓解生物和非生物胁迫。沙雷氏菌的各种农业应用包括直接使用细菌细胞进行种子包衣、叶面喷洒和土壤接种,或将其生物活性化合物单独或与其他材料结合应用于植物的各个部位。这些努力旨在增强植物健康、抑制疾病和控制害虫种群。尽管应用前景广阔,但有报道称植物和动物具有机会性致病性。因此,应考虑几种安全方法和使用毒力因子突变菌株。沙雷氏菌在农业中的应用趋势预计将持续下去。

UDP-糖基转移酶是决定广谱和专效灰翅夜蛾宿主植物范围的关键因素
植食性昆虫已经进化出复杂的解毒系统来克服许多植物产生的抗食草动物化学防御。然而,这些生物转化系统在通才和专才昆虫物种中有何不同,以及它们在确定昆虫宿主植物范围方面的作用仍是一个悬而未决的问题。在这里,我们表明 UDP - 葡萄糖基转移酶 (UGT) 在确定 Spodoptera 属内昆虫物种的宿主范围方面起着关键作用。对宿主植物宽度不同的 Spodoptera 物种进行比较基因组分析,发现在通才物种中 UGT 基因数量相对保守,但在专才 Spodoptera picta 中 UGT 基因假基因化水平较高。CRISPR - Cas9 敲除 Spodoptera frugiperda 的三个主要 UGT 基因簇表明,UGT33 基因在使该物种利用禾本科植物玉米、小麦和水稻方面发挥重要作用,而 UGT40 基因促进棉花的利用。进一步的体内和体外功能分析表明,UGT SfUGT33F32 是使广谱 S. frugiperda 能够解毒苯并恶嗪类化合物 DIMBOA(2,4-二羟基-7-甲氧基-2H-1,4-苯并恶嗪-3(4H)-酮)的关键机制,DIMBOA 是由禾本科植物产生的强效杀虫毒素。然而,虽然这种解毒能力在几种广谱 Spodoptera 物种中得到了保留,但专食文殊兰植物的 Spodoptera picta 因 SpUGT33F34 的非功能性突变而无法解毒 DIMBOA。总之,这些发现为了解昆虫 UGT 在宿主植物适应中的作用、广谱和专谱之间进化转变的机制基础提供了见解,并为控制一组臭名昭著的害虫提供了分子目标。

非洲绿松石Killifish(Nothobranchius Furzeri)
有1200多种杀虫物种,除澳大利亚和南极洲外,几乎每个大陆都有大陆。大多数杀伤力都是小的鱼,平均尺寸为2.5 e 7 cm,可以大致分为短寿命的年度物种和寿命更长的非年轻人。killifhs是Cyprinodontiform Order的成员,也称为“牙齿”,这是一个包括几个家庭(例如Aplocheilidae,Nothobranchiidae,Cyprinodontidae,Cyprinodontidae,endululidae,其他)(Nelson等人)(Nelson等人(Nelson等,2016; Scheel,1990年))。近年来,由于其自然压缩的寿命,快速的性成熟度,胚胎性渗透,胚胎渗透速度快速(在干旱季节的生存机制),以及在干旱季节的短期和2015年,2015年,2015; Et et et et e et et e et et et et et e et et e et eet et et e et eet et et e et et et e et et e et eet et et e et et et e et et e et et e et et e et et et e et et e et eet et eet et et et e et eet et eet et et e et et e et et e et e et。 Al。,2016年; Platzer和Englert,2016年)11.1)。这些生活历史特征可能是适应其季节性栖息地,东南非洲的短暂的水锅,主要是津巴布韦和莫桑比克,那里只有在短暂的雨季期间可用的水(Cellerino等,2015; Harel and Brunet; Harel and Brunet,2015; Kim等,2016; Kim等,2016; Platser and Enger and Enger,2016年)11.2a)。1968年在贡纳雷佐国家公园(津巴布韦)收集了绿松石的杀戮鱼。以公园g ona r e z khou的名字命名的近交Grz菌株(在当地的Shona方言中为“大象的位置”)目前是最多的

农场内生产微生物昆虫病原体以供使用……
摘要 在巴西,种植者生产有益微生物专供自己使用是一种被称为“农场生产”的做法。至于农场生物杀虫剂,它们最初在 20 世纪 70 年代用于防治多年生和半多年生作物的害虫,但自 2013 年以来,其使用范围已扩大到玉米、棉花和大豆等一年生作物的害虫。目前,数百万公顷的土地正在使用这些农场制剂进行处理。本地生产可降低成本、满足当地需求并减少对环境有害的化学农药的投入,从而有助于建立更可持续的农业生态系统。批评人士认为,如果不实施严格的质量控制措施,农场制剂可能会:(1) 被可能包括人类病原体的微生物污染,或 (2) 含有极少的活性成分,影响田间药效。细菌杀虫剂的农场发酵占主导地位,尤其是针对鳞翅目害虫的苏云金芽孢杆菌。然而,在过去的 5 年中,昆虫病原真菌的生产迅速增长,主要用于控制吸食汁液的昆虫,例如粉虱(Bemisia tabaci (Gennadius))和玉米叶蝉(Dalbulus maidis (DeLong and Wolcott))。相比之下,昆虫病毒的农场生产增长有限。巴西约 500 万农村生产者中的大多数拥有中小型农场,虽然绝大多数人仍然没有在农场生产生物农药,但这个话题已经引起了他们的兴趣。许多采用这种做法的种植者通常使用非无菌容器作为发酵罐,导致制剂质量差,并且有失败的案例报道。另一方面,一些非正式报告表明,即使受到污染,农场制剂也可能有效,这至少可以部分解释为液体培养基中的微生物池分泌的杀虫次生代谢物。事实上,关于这些微生物生物农药的功效和作用方式的信息不足。通常是大型农场生产的生物农药污染程度较低,其中一些农场的连续耕地面积超过 20,000 公顷,因为其中许多农场拥有先进的生产设施,并拥有专业知识和训练有素的员工。农场生物农药的使用预计将持续下去,但采用率将取决于多种因素,例如选择安全、毒性强的微生物菌株和实施合理的质量控制措施(符合新兴的巴西法规和国际标准)。本文介绍并讨论了农场生物杀虫剂的挑战和机遇。
全球基金在肯尼亚共和国的资助
尽管经历了一段艰难和充满挑战的时期,肯尼亚在抗击这三种疾病方面取得了进展。艾滋病毒感染率从 2018 年的 4.9% 下降到 2020 年的 4.5%,艾滋病毒发病率从 2016 年的 0.27% 下降到 2020 年的 0.14%。结核病防治也取得了进展,2018 年至 2020 年期间发病率下降了 11%。然而,进一步的进展受到严重阻碍,因为流程效率低下,延误了采购,影响了整个供应链的药品供应。虽然肯尼亚医疗用品管理局 (KEMSA) 1 正在以有竞争力的价格采购质量有保证的商品,但周转时间过长导致延误,损害了项目。仓储和配送控制不力,影响了数据质量和商品可追溯性,以及中央和医疗机构层面的药品供应。没有一个强大的系统来监测、跟踪和报告运送到医疗机构的商品,无法完全说明收到的药品。采购和供应链管理系统被评为无效。尽管 2020 年和 2021 年 COVID-19 严重扰乱了全球基金项目,但赠款大多表现良好,关键影响指标呈积极趋势。2020 年,150 万艾滋病毒感染者中约有 96% 知道自己的感染状况,89% 正在接受治疗,94% 的病毒载量得到抑制。2 过去 10 年,新感染病例减少了 44%。疟疾患病率和结核病发病率都有所下降。然而,并非所有项目方面都步入正轨。由于 COVID 限制而做出的决定推迟了计划于 2020 年进行的长效杀虫蚊帐分发活动,这可能会增加疾病的患病率、发病率和死亡率。如果不能减少漏诊的结核病病例数量,将会增加疾病的发病率和患病率。肯尼亚的项目干预措施被评为部分有效。肯尼亚及时吸收新冠资金的能力仍然很低,在拨款结束时仅为 51%,这主要是由于采购拖延所致。资金利用率低限制了该国快速应对疫情的能力。新冠资金的管理,包括拨款灵活性,需要显著改进。

安妮·奥弗斯特里特
Anne Overstreet 生物农药和污染防治部(7511P) 环境保护署农药计划办公室 1200 Pennsylvania Ave. NW 华盛顿特区 20460–0001 事由:卷宗编号 EPA–HQ–OPP–2019–0508 2020 年 12 月 8 日 我谨代表下列农民、牧场主、合作社、零售商、科学家、植物育种者、种子生产者和共同监管者,代表美国广泛而多样的农业利益相关者,感谢有机会就拟议规则“农药;源自新技术的某些植物内保护剂 (PIP) 的豁免”发表评论和反馈意见。我们赞扬美国环境保护署 (EPA) 为实现生物技术监管体系现代化而做出的努力,该署提议将符合条件的“基于通过生物技术创造的性相容植物的 PIP”从《联邦杀虫剂、杀菌剂和灭鼠剂法案》 (FIFRA) 的大部分要求以及《联邦食品药品和化妆品法案》 (FFDCA) 规定的容差设定要求中豁免。我们赞赏拟议规则的总体愿景,但我们也提出了一些建议,我们认为这些建议将有助于 EPA 制定更科学、更基于风险的最终规则。我们还相信,如果这些建议被采纳,将有助于美国保持其在植物生物技术发展方面的全球领导地位。我们在此解释了统一的基线建议,以增强拟议规则,满足我们各利益相关者的需求。许多签署方还将提交单独的意见,提供与各个利益相关者需求相关的具体建议的更多细节,或提出超出本信函内容的建议。拟议的 PIP 豁免的近期历史背景 了解促使 EPA 提议豁免这一范围狭窄、风险较低的 PIP 子集的近期历史背景非常重要。2015 年 7 月,奥巴马总统的总统行政办公室 (EOP) 发布了一份备忘录,提出了对当前生物技术监管框架在某些情况下强加不必要的成本和负担的担忧,这些成本和负担阻碍了中小企业参与市场,限制了公众对监管流程的理解,从本质上抑制了创新。1 该备忘录成立了一个跨部门工作组,以制定“现代化生物技术产品监管体系的国家战略”(国家战略),该战略于 2016 年 9 月发布。除了重申“美国政府的政策是寻求保护健康和环境的监管方法,同时减少监管负担,避免不合理地抑制创新、污名化新技术或制造贸易壁垒”之外,国家战略还指示 EPA 应该“阐明其对源自基因组编辑技术的杀虫产品的态度。”2
NIC-2004_detail_19jan2009.pdf - MCA
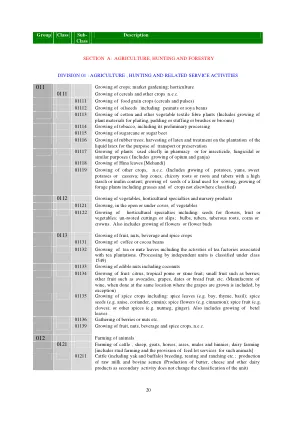
011 作物种植;市场园艺;园艺 0111 谷物和其他未另分类作物的种植01111 粮食作物(谷物和豆类)的种植 01112 油籽(包括花生或大豆)的种植 01113 棉花和其他植物纺织纤维植物的种植(包括用于编织、衬垫或填料或刷子或扫帚的植物材料的种植) 01114 烟草的种植,包括其初加工 01115 甘蔗或甜菜的种植 01116 橡胶树的种植;收获乳胶并在种植园中对液态乳胶进行处理以供运输或保存 01117 种植主要用于制药或杀虫、杀菌或类似用途的植物(包括种植鸦片和大麻) 01118 种植 Hina 叶 [Mehandi] 01119 种植其他未列明的作物(包括种植土豆、山药、红薯或木薯;啤酒花球果、菊苣根或含有高淀粉或菊粉的根和块茎;种植用于播种的种子,种植包括草在内的饲料植物以及未分类的作物) 0112 种植蔬菜、园艺特产和苗圃产品 01121 在露天或有遮盖的情况下种植蔬菜 01122 种植园艺特产,包括:花卉、水果或蔬菜种子;无根插枝或接穗;球茎、块茎、块根、玉米或冠。还包括花卉或花蕾的种植 0113 水果、坚果、饮料和香料作物的种植 01131 咖啡豆或可可豆的种植 01132 茶叶或马黛茶叶的种植,包括与茶园相关的茶厂活动。(独立单位的加工归类为 1549 类) 01133 食用坚果的种植,包括椰子 01134 水果的种植:柑橘、热带仁果或核果;小果实,如浆果;其他水果,如鳄梨、葡萄、枣或面包果等。(葡萄酒的制造,在葡萄生长的同一地点进行,但例外) 01135 香料作物的种植,包括:香料叶(例如月桂、百里香、罗勒);香料种子(例如茴香、芫荽、小茴香);香料花(例如肉桂);香料果实(例如丁香);或其他香料(例如肉豆蔻、生姜)。还包括槟榔叶的种植。01136 浆果或坚果等的采集01139 水果、坚果、饮料和香料作物的种植,未另分类;生牛奶和牛精液的生产(生产黄油、奶酪和其他乳制品作为次要活动不会改变单位的分类)012 动物养殖 0121 牛、羊、山羊、马、驴、骡和驴驹的养殖;奶牛养殖[包括种马养殖和为此类动物提供饲养场服务] 01211 牛(包括牦牛和水牛)的繁殖、饲养和放牧等

苏云金芽孢杆菌及其毒素的生物技术进步:最近的更新
广泛的害虫,主要是鳞翅目(毛毛虫),双翅目(蚊子和黑蝇)和鞘翅目(甲虫幼虫)(Sanchis 2011)。bt的特征是在孢子形成过程中生产,内毒素蛋白(称为哭泣的蛋白),这些蛋白会积聚并形成晶体包含体。昆虫必须消耗/摄取这些哭泣的蛋白质,才能感受到其作用,直到昆虫死亡。在摄入后,昆虫中肠内的碱性条件会导致晶体的溶解化,从而将其转化为有毒的核心碎片(Sansinenea 2019)。这些有毒蛋白与位于昆虫中肠上皮细胞上的受体(糖蛋白或糖蛋白)结合(Bravo等人2011)。结合后,毒素会改变其构象,从而使其插入细胞膜并形成阳离子选择通道(Bravo等。2013)。当形成足够的这些通道时,几个阳离子进入了细胞。这会导致细胞内部的渗透不平衡,从而导致中肠上皮完整性的丧失。这使碱性肠道果汁和细菌可以通过中肠地下膜,杀死昆虫。当用作喷雾剂时,这些毒素无效地防止昆虫攻击植物的根或植物的内部部分(Sanahuja等人。2011)。这些局限性引发了人们对开发新的遗传修饰植物和细菌表达哭泣和其他BT-杀虫基因的兴趣,以便提供更有效的毒素递送系统来控制这些昆虫(Azizoglu和Karabörklü2021)。2021; Lazarte等。在生物技术技术(例如基因工程)中的持续进展,具有计算生物学的能力,导致了有关BT的发展和发现。在这种情况下,全球各个研究小组对寻找具有新的抑制活性范围和高水平的毒性毒素的新型哭泣毒素非常感兴趣,这是针对虫害的一种替代品,这种毒性毒性具有更高的抗药性水平(Hou等人 2019; Crickmore等。 2021)。 结果,使用术基因组数据,遗传修饰(GM)微生物的发展的持续菌株改善正在成为不可避免的能够实现非本地基因表达和改善本机生产国以发展遗传学改善菌株的工具包(Liu等人(Liu等)(Liu等人。 2017; Azizoglu等。 2020)。 今天的新一代方法,例如模拟和动态研究,2019; Crickmore等。2021)。结果,使用术基因组数据,遗传修饰(GM)微生物的发展的持续菌株改善正在成为不可避免的能够实现非本地基因表达和改善本机生产国以发展遗传学改善菌株的工具包(Liu等人(Liu等)(Liu等人。2017; Azizoglu等。2020)。今天的新一代方法,例如模拟和动态研究,

Shane Denecke
1。Vorgia E.,M。Lamprousi,S。Denecke,K。Vogelsang,S。Geibel等,2021年的功能特征和转录组中的中腹细胞系中的中腹细胞系(Lep-Idoptera:noctuidae)。昆虫生物化学。mol。生物。128:103510。https://doi.org/10.1016/j.ibmb.2020.103510 2。Swevers L.,S。Denecke,K。Vogelsang,S。Geibel和J. Vontas,2020年,哺乳动物类器官技术可以应用于昆虫肠道吗?害虫管理。SCI。 77:55–63。 https://doi.org/10.1002/ps.6067 3。 DENECKE S.* M.,O。DRIVA,H。N. B. Luong,P。Ioannidis,M。Linka等,2020年,溶质载体超家族在节肢动物中的识别和进化趋势。 基因组生物。 Evol。 12:1429–1439。 https://doi.org/10.1093/gbe/evaa153 4。 Samantsidis G.-R.,R。Panteleri,S。Denecke,S。Kounadi,I。Christou等,2020年,“我无法创造的东西,我不理解”:在功能验证的代谢和目标位点昆虫抗药性的协同作用。 proc。 R. Soc。 B Biol。 SCI。 287:20200838。https://doi.org/10.1098/rspb.2020.0838 5。 douris V.,S。Denecke,T。VanLeeuwen,C。Bass,R。Nauen等,2020年,使用CRISPR/CAS9基因组修饰来理解杀虫剂耐药性的遗传基础:果蝇及以后。 农药。 生物化学。 生理学。 167。https://doi.org/10.1016/j.pestbp.2020.1045956。 昆虫分子。 生物。 29:363–372。 https://doi.org/10.1111/imb.12640 7。SCI。77:55–63。https://doi.org/10.1002/ps.6067 3。 DENECKE S.* M.,O。DRIVA,H。N. B. Luong,P。Ioannidis,M。Linka等,2020年,溶质载体超家族在节肢动物中的识别和进化趋势。 基因组生物。 Evol。 12:1429–1439。 https://doi.org/10.1093/gbe/evaa153 4。 Samantsidis G.-R.,R。Panteleri,S。Denecke,S。Kounadi,I。Christou等,2020年,“我无法创造的东西,我不理解”:在功能验证的代谢和目标位点昆虫抗药性的协同作用。 proc。 R. Soc。 B Biol。 SCI。 287:20200838。https://doi.org/10.1098/rspb.2020.0838 5。 douris V.,S。Denecke,T。VanLeeuwen,C。Bass,R。Nauen等,2020年,使用CRISPR/CAS9基因组修饰来理解杀虫剂耐药性的遗传基础:果蝇及以后。 农药。 生物化学。 生理学。 167。https://doi.org/10.1016/j.pestbp.2020.1045956。 昆虫分子。 生物。 29:363–372。 https://doi.org/10.1111/imb.12640 7。https://doi.org/10.1002/ps.6067 3。DENECKE S.* M.,O。DRIVA,H。N. B. Luong,P。Ioannidis,M。Linka等,2020年,溶质载体超家族在节肢动物中的识别和进化趋势。基因组生物。Evol。12:1429–1439。https://doi.org/10.1093/gbe/evaa153 4。Samantsidis G.-R.,R。Panteleri,S。Denecke,S。Kounadi,I。Christou等,2020年,“我无法创造的东西,我不理解”:在功能验证的代谢和目标位点昆虫抗药性的协同作用。proc。R. Soc。 B Biol。 SCI。 287:20200838。https://doi.org/10.1098/rspb.2020.0838 5。 douris V.,S。Denecke,T。VanLeeuwen,C。Bass,R。Nauen等,2020年,使用CRISPR/CAS9基因组修饰来理解杀虫剂耐药性的遗传基础:果蝇及以后。 农药。 生物化学。 生理学。 167。https://doi.org/10.1016/j.pestbp.2020.1045956。 昆虫分子。 生物。 29:363–372。 https://doi.org/10.1111/imb.12640 7。R. Soc。B Biol。 SCI。 287:20200838。https://doi.org/10.1098/rspb.2020.0838 5。 douris V.,S。Denecke,T。VanLeeuwen,C。Bass,R。Nauen等,2020年,使用CRISPR/CAS9基因组修饰来理解杀虫剂耐药性的遗传基础:果蝇及以后。 农药。 生物化学。 生理学。 167。https://doi.org/10.1016/j.pestbp.2020.1045956。 昆虫分子。 生物。 29:363–372。 https://doi.org/10.1111/imb.12640 7。B Biol。SCI。 287:20200838。https://doi.org/10.1098/rspb.2020.0838 5。 douris V.,S。Denecke,T。VanLeeuwen,C。Bass,R。Nauen等,2020年,使用CRISPR/CAS9基因组修饰来理解杀虫剂耐药性的遗传基础:果蝇及以后。 农药。 生物化学。 生理学。 167。https://doi.org/10.1016/j.pestbp.2020.1045956。 昆虫分子。 生物。 29:363–372。 https://doi.org/10.1111/imb.12640 7。SCI。287:20200838。https://doi.org/10.1098/rspb.2020.0838 5。douris V.,S。Denecke,T。VanLeeuwen,C。Bass,R。Nauen等,2020年,使用CRISPR/CAS9基因组修饰来理解杀虫剂耐药性的遗传基础:果蝇及以后。农药。生物化学。生理学。167。https://doi.org/10.1016/j.pestbp.2020.1045956。昆虫分子。生物。29:363–372。https://doi.org/10.1111/imb.12640 7。Koidou V.,S。Denecke*,P。Ioannidis,I。Vlatakis,I。Livadaras等,2020年,有效的CRISPR/CAS9介导的基因组介导的基因组编辑。denecke s*。,P。ioannidis*,B。Buer,A。Ilias,V。Douris等,2020年,Nezara Viridula(杂翅目:五翅目:pentatomidae)中表达的转录组和蛋白质组学图,Midgut提出了心苯基植物的分类性,并表明了心齿植物的分类。BMC基因组学21:129。https://doi.org/10.1186/S12864-020-6459-6 8。Riga M.,S。Denecke*,I。Livadaras,S。Geibel,R。Nauen等,2020年,在Nezara viridula中开发有效RNAi,用于杀虫剂靶标。拱门。昆虫生物化学。生理学。103:E21650。 https://doi.org/10.1002/arch.21650 9。 Young H. K.,S。M. Denecke,C。Robin和A. Fournier级,2019年,幼虫暴露于咪二藻中的幼虫会影响果蝇中的成人行为。 J. Evol。 生物。 33:151–164。 https://doi.org/10.1111/jeb.13555 10。 Denecke S*。,L。Swevers,V。Douris和J. Vontas,2018年,口腔杀虫化合物如何穿越昆虫中肠上皮? 昆虫生物化学。 mol。 生物。 103:22–35。 https://doi.org/10.1016/ j.ibmb.2018.10.005 11。 harrop T. W.r§。,S。Denecke§,Y。T。Yang,J。Chan,P。J。Daborn等,2018,通过果蝇中的线粒体细胞色素P450激活Nitenpyram的证据。 害虫管理。 SCI。 74:1616–1622。 https://doi.org/10.1002/ps.4852 12. 昆虫生物化学。 mol。103:E21650。https://doi.org/10.1002/arch.21650 9。Young H. K.,S。M. Denecke,C。Robin和A. Fournier级,2019年,幼虫暴露于咪二藻中的幼虫会影响果蝇中的成人行为。J. Evol。 生物。 33:151–164。 https://doi.org/10.1111/jeb.13555 10。 Denecke S*。,L。Swevers,V。Douris和J. Vontas,2018年,口腔杀虫化合物如何穿越昆虫中肠上皮? 昆虫生物化学。 mol。 生物。 103:22–35。 https://doi.org/10.1016/ j.ibmb.2018.10.005 11。 harrop T. W.r§。,S。Denecke§,Y。T。Yang,J。Chan,P。J。Daborn等,2018,通过果蝇中的线粒体细胞色素P450激活Nitenpyram的证据。 害虫管理。 SCI。 74:1616–1622。 https://doi.org/10.1002/ps.4852 12. 昆虫生物化学。 mol。J. Evol。生物。33:151–164。https://doi.org/10.1111/jeb.13555 10。Denecke S*。,L。Swevers,V。Douris和J. Vontas,2018年,口腔杀虫化合物如何穿越昆虫中肠上皮?昆虫生物化学。mol。生物。103:22–35。https://doi.org/10.1016/ j.ibmb.2018.10.005 11。harrop T. W.r§。,S。Denecke§,Y。T。Yang,J。Chan,P。J。Daborn等,2018,通过果蝇中的线粒体细胞色素P450激活Nitenpyram的证据。害虫管理。SCI。 74:1616–1622。 https://doi.org/10.1002/ps.4852 12. 昆虫生物化学。 mol。SCI。74:1616–1622。https://doi.org/10.1002/ps.4852 12. 昆虫生物化学。 mol。https://doi.org/10.1002/ps.4852 12.昆虫生物化学。mol。denecke s。,R。Fusetto和P. Batterham,2017年,使用CRISPR-CAS9敲除果蝇Melanogaster ABC转运蛋白在杀虫剂生物学中的作用。生物。91:1-9。 https://doi.org/10.1016/j.ibmb.2017.09.017 13。 DeNecke S.,R。Fusetto,F。Martelli,A。Giang,P。Battlay等,2017,2017年多个P450和神经元基因的变化,这是对果蝇大众群中对杀虫剂咪二酸的反应。 SCI。 Rep。7:11338。https://doi.org/10.1038/S41598-017-11092-5 14。 Fusetto R.,S。Denecke,T。Perry,R。A。J. O'Hair和P. Batterham,2017年,将CYP6G1和肠道微生物在果蝇中杀虫剂咪二氯吡啶的代谢中的作用分开。 SCI。 Rep。7:11339。https://doi.org/10.1038/S41598-017-09800-2 15。 DeNecke S.,C。J. Nowell,A。Fournier级,T。Perry和P. Batterham,2015年Wiggle索引:一种开源生物测定,用于评估果蝇中果蝇中的亚致死性杀虫剂反应。 PLOS ONE 10:E0145051。 https://doi.org/10.1371/journal.pone.014505191:1-9。https://doi.org/10.1016/j.ibmb.2017.09.017 13。DeNecke S.,R。Fusetto,F。Martelli,A。Giang,P。Battlay等,2017,2017年多个P450和神经元基因的变化,这是对果蝇大众群中对杀虫剂咪二酸的反应。SCI。 Rep。7:11338。https://doi.org/10.1038/S41598-017-11092-5 14。 Fusetto R.,S。Denecke,T。Perry,R。A。J. O'Hair和P. Batterham,2017年,将CYP6G1和肠道微生物在果蝇中杀虫剂咪二氯吡啶的代谢中的作用分开。 SCI。 Rep。7:11339。https://doi.org/10.1038/S41598-017-09800-2 15。 DeNecke S.,C。J. Nowell,A。Fournier级,T。Perry和P. Batterham,2015年Wiggle索引:一种开源生物测定,用于评估果蝇中果蝇中的亚致死性杀虫剂反应。 PLOS ONE 10:E0145051。 https://doi.org/10.1371/journal.pone.0145051SCI。Rep。7:11338。https://doi.org/10.1038/S41598-017-11092-5 14。 Fusetto R.,S。Denecke,T。Perry,R。A。J. O'Hair和P. Batterham,2017年,将CYP6G1和肠道微生物在果蝇中杀虫剂咪二氯吡啶的代谢中的作用分开。 SCI。 Rep。7:11339。https://doi.org/10.1038/S41598-017-09800-2 15。 DeNecke S.,C。J. Nowell,A。Fournier级,T。Perry和P. Batterham,2015年Wiggle索引:一种开源生物测定,用于评估果蝇中果蝇中的亚致死性杀虫剂反应。 PLOS ONE 10:E0145051。 https://doi.org/10.1371/journal.pone.0145051Rep。7:11338。https://doi.org/10.1038/S41598-017-11092-5 14。Fusetto R.,S。Denecke,T。Perry,R。A。J. O'Hair和P. Batterham,2017年,将CYP6G1和肠道微生物在果蝇中杀虫剂咪二氯吡啶的代谢中的作用分开。SCI。 Rep。7:11339。https://doi.org/10.1038/S41598-017-09800-2 15。 DeNecke S.,C。J. Nowell,A。Fournier级,T。Perry和P. Batterham,2015年Wiggle索引:一种开源生物测定,用于评估果蝇中果蝇中的亚致死性杀虫剂反应。 PLOS ONE 10:E0145051。 https://doi.org/10.1371/journal.pone.0145051SCI。Rep。7:11339。https://doi.org/10.1038/S41598-017-09800-2 15。 DeNecke S.,C。J. Nowell,A。Fournier级,T。Perry和P. Batterham,2015年Wiggle索引:一种开源生物测定,用于评估果蝇中果蝇中的亚致死性杀虫剂反应。 PLOS ONE 10:E0145051。 https://doi.org/10.1371/journal.pone.0145051Rep。7:11339。https://doi.org/10.1038/S41598-017-09800-2 15。DeNecke S.,C。J. Nowell,A。Fournier级,T。Perry和P. Batterham,2015年Wiggle索引:一种开源生物测定,用于评估果蝇中果蝇中的亚致死性杀虫剂反应。PLOS ONE 10:E0145051。https://doi.org/10.1371/journal.pone.0145051