XiaoMi-AI文件搜索系统
World File Search System极性相反

使用单克隆抗体分析粘液虫(粘菌门:粘孢子门)孢子抗原
摘要:开发了一种采用 Percoll™ 梯度离心法从大西洋鲑 Salmo salar 的体肌组织中纯化 Kudoa thyrsites 孢子的方法。然后用高度纯化的孢子免疫近交系 BALB/c 小鼠,以衍生分泌 Kudoa 特异性单克隆抗体 (mAb) 的杂交瘤。通过免疫荧光显微镜和流式细胞术对 mAb 进行分析表明,几种 mAb 对 K. thyrsites 孢子表面的抗原具有特异性,而其他 mAb 与 K. thyrsites、K. paniformis 和 K. crumena 孢子的极性荚膜或极性细丝发生反应。使用表面结合 mAb 对孢子裂解物进行免疫印迹,结果显示 46 至 >220 kDa 的宽条带,而针对极性荚膜和极性细丝抗原的特异性 mAb 检测到不同分子量的更清晰条带,具体取决于 Kudoa 物种。K. thyrsites 孢子表面抗原的主要表位被证明是碳水化合物,这是由其对无水三氟甲烷磺酸处理的敏感性和对蛋白酶 K 处理的抗性决定的。使用 K. thyrsites 特异性 mAb 对分离的、完整的、透化的疟原虫和含有疟原虫的体细胞肌肉组织薄切片进行免疫荧光显微镜检查,发现在产生孢子的疟原虫和受感染的大西洋鲑鱼肉中都有孢子的强烈标记。通过免疫印迹法检测到的孢子只有 100 个,表明这些 mAb 具有用于开发基于现场的诊断测试的潜力。

新合成的抗肿瘤药物候选物2H4MBBH与人血清白蛋白相互作用的光谱和热力学表征
不同的氟、羟基和甲氧基取代的苯甲醛残基(图 1)对分离的螺旋体肌肉幼虫表现出显着的体外驱虫活性,以及对 MCF-7 和 AR-230 乳腺癌细胞的强效抗增殖活性(Anichina 等人 2021;Argirova 等人 2021、2023)。这些化合物还能够抑制微管蛋白聚合(Argirova 等人 2021)。含有羟基苯基和甲氧基苯基部分的 1H-苯并咪唑-2-基腙在卵磷脂和脱氧核糖模型系统中表现出强大的抗氧化和自由基清除特性以及铁诱导的氧化损伤。密度泛函理论计算表明,1H-苯并咪唑-2-基腙具有非常通用的自由基清除特性,这是因为存在多个反应位点,这些反应位点的特点是反应焓相对较低,并且可以通过不同的反应途径同时起作用:非极性介质中的氢原子转移、极性介质中的连续质子损失电子转移以及极性和非极性介质中的自由基加合物形成 (Argirova 等人,2021 年)。我们选择在这里检查化合物 2H4MB-BH 施加后 HSA 的荧光曲线,并利用这些曲线表征 2H4MBBH-HSA 相互作用参数。所采取的方法是表明荧光参数有显著的变化,这将有助于评估合成的抗癌镇静剂 2-(2-羟基-4-甲氧基苄亚甲基)-1-(1H-苯并咪唑-2-基)肼的恢复效果。
Annamacharya技术与科学学院
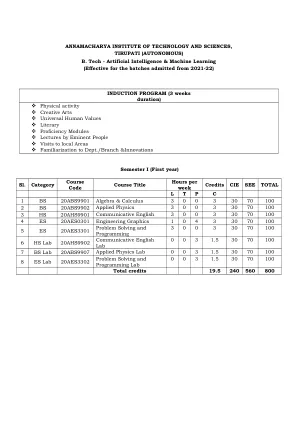
(自治)人工智能(AI)年:I学期:I研究分支:AIML课程代码年度和SEM代数和计算L T P C 20ABS9901 I-I 3 0 0 3课程成果:在学习课程后,学生将能够Co1。将矩阵代数技术应用于求解各种线性方程。二氧化碳。分析二次形式和平均值定理的线性变换。二氧化碳。将部分导数的基本概念应用于多变量函数。CO4。 评估笛卡尔,极性,圆柱和球形坐标的多个积分CO4。评估笛卡尔,极性,圆柱和球形坐标的多个积分

铁弹性域壁内的抗fiferroelectric Switch
图2在室温下(t = 300k),在正骨catio 3中(110)型DWS的结构和极性特性。(a)(110)dw的几何图形和在catio 3的正栓相中的几何学和方向的草图。(b)是由两个平行DWs组成的三明治模型,具有反平行DW极向量(绿色箭头)。DW内部的铁弹性双角和最大极化为C.A.0.52和2.4c/cm -2。插图(b)是通过透射电子显微镜(TEM)获得的DW内部的极向量[16]。X-Y,X-Z和Y-Z(双壁平面)平面内DW极化的局部细节显示在(C-D),(E-F)和(G-H)中。绿色和红色箭头是与图相对应的奇数甚至层的极性向量。1(d)。小极化倾斜存在于X-Y和X-Z平面内,而在双壁(Y-Z)内发现了相对较大的倾斜度。由于全球倒置中心对称性的保护,附近双壁的总体极化向量取消了。极性向量箭头被放大150倍以进行澄清。
![arxiv:2412.18878v1 [physics.optics] 2024年12月25日](/simg/5\59b14591d8d8c891d6dd044e5a15904dbdc546f6.webp)
arxiv:2412.18878v1 [physics.optics] 2024年12月25日
传统的金属和N型掺杂的半导体材料是新兴的Epsilon – Near -near -Zero(ENZ)材料,展示了非线性光子应用的巨大潜力。然而,这种材料的一个重要限制是缺乏多功能的ENZ波长调整,因此,对ENZ波长的动态调整仍然是一个技术挑战,从而限制了其潜在应用,例如多频带通信。在此,通过光激发后极性形成/解耦的孔浓度的可逆变化以及可调的enz波长移动,可以观察到PSS膜中的ENZ波长的动态调整,从而可以通过可逆的孔浓度变化来实现PSS膜。关于极性激发超快速动力学的实验研究表明,北极子积累的〜80 fs时间常数,北极子解耦的〜280 fs时间常数,表明在子picosocecond时尺度内的enz波长逆转超快切换的潜力。这些发现表明,P型有机半导体可以用作通过极性激发动态调整ENZ波长的新型平台,这为基于ENZ的非线性光学应用在柔性光电上开辟了新的可能性。

通过体细胞核转移(SCNT)
这种SCNT卵母细胞的人工激活导致细胞分裂和染色体分离为伪极性体,并以70%的效率下的二核原体。与正常二倍体(n = 46)数量相比,极性体和Zygotes中单个染色体的下一代测序表明,染色体的数量降低了近一半(n = 19)(n = 19)。同源对的全面测序表明,平均将23对同源对的一半(n = 11)正确分离为极体和合子,而剩余的染色体对保持在一起,导致了肾上变。未检测到体细胞同源物之间的重组证据。

随机饱和诱变生成高度多样化的定向进化文库
为了了解每种野生型氨基酸对不同侧链性质的可及性,我们将所有 20 种氨基酸分为 8 类:非极性(NP、M、I、L、V、A)、极性不带电(PU、S、T、Q、N)、带正电荷(PC、R、K、L)、带负电荷(NC、D、E)、芳香族(Ar、F、T、Y)和三个特殊基团 P、C、G,由于其性质不同,每个基团仅由一个氨基酸组成。通过易错 PCR,每种野生型氨基酸都有一些不可接近的性质类别,如图 4c 所示。此外,在

存储反铁电PbZrO3
基于反铁电的介电电容器因其出色的储能性能和在收集脉冲功率方面的非凡灵活性而备受关注。尽管如此,迄今为止,尚未阐明与储能过程固有耦合的原位原子级结构演化途径,以最终理解其机制。本文报道了反铁电PbZrO 3 在存储电子束照射的能量过程中的时间和原子分辨率结构相演变。通过采用最先进的负球差成像技术,本文介绍的定量透射电子显微镜研究阐明了与晶胞体积变化和极化旋转相关的极性氧八面体的层次演化解释了逐步的反铁电到铁电相变。特别是,在动态结构研究过程中建立了一种非常规的铁电类别——具有独特摆线极化序的铁电畸变相。通过阐明原子尺度相变途径,该研究的结果为探索具有非极性到极性相变的储能材料中的新型铁致畸变相开辟了一个新领域。

极地设计公司为太空应用设计了抗辐射 14T SRAM 单元
摘要 本文提出了一种用于航天应用的抗辐射极性设计 14T (RHPD-14T) SRAM 单元。通过估算 65 纳米互补金属氧化物半导体 (CMOS) 技术的各种设计指标,分析了所提出的 RHPD-14T 单元的性能。基于结合抗辐射极性设计技术与合理的布局拓扑,所提出的 RHPD-14T 可以耐受所有单节点翻转和部分双节点翻转。仿真结果表明,RHPD-14T 的写入访问时间比 RSP-14T/QUCCE-10T/DICE/S4P8N/We-Quatro(@VDD=1.2V) 短 1.83 倍 / 1.59 倍 / 1.56 倍 / 1.12 倍 / 1.05 倍。 RHPD-14T的字线写触发电压比QUCCE-10T/DICE/We-Quatro/S4P8N/RSP-14T (@VDD=1.2V)高2.67×/2.22×/1.35×/1.29×/1.26×;RHPD-14T的保持静态噪声容限比DICE/S4P8N/RHPD-12T (@VDD=1.2 V)高14.85×/7.15×/1.05×。此外,蒙特卡洛(MC)模拟证明RHPD-14T波动性小、稳定性强、恢复能力稳定、抗单效应翻转(SEU)能力强。关键词:保持静态噪声容限、极性设计抗辐射、单效应翻转分类:集成电路

使用智能设备探索磁性
外部封面:为Helmholtz Coil设置的实验(信用:Sten Odenwald);电磁阀模型(信用:Paul Nylander);进行极性测量设置(信用:Sten Odenwald);智能手机显示(信用:Sten Odenwald);黑子上的磁性环(信用:NASA/TRACE)。内部封面:从左到右的顶行:太阳上的磁性线(NASA/SDO);笛卡尔的力线草图;地球磁场模型(信用:Gary A. Glatzmaier-洛斯阿拉莫斯国家实验室 - 美国能源部)。从左到右的底行:大强子(信用:CERN);磁性指南针的方向(信用:NOAA);黑子极性地图(信用:NASA/SDO)