XiaoMi-AI文件搜索系统
World File Search System看到

看到的是:可视化工具箱,以提高玉米基因组编辑的选择效率
CRISPR/CAS9基因编辑的新兴和有希望的生物技术方法正在彻底改变作物的改善。然而,在转化或之后或之后的正面选择,较低的及时性和劳动力的性质以及随后对突变的识别是其农业应用的主要挑战,是从上游(高发射)突变体筛选到下游商业商业生产(He and Zhao,Zhao,2020)。尽管已使用视觉标记,尤其是包括绿色荧光蛋白(GFP)和红色荧光蛋白(RFP)的荧光标记,已被用于快速可视化转基因材料(Qi等人,2020年)(2020年)(2020年),但成本效益和非侵蚀性crispr-secr-dectr-decration and Crist-crist crist-crist-defty Gene and noff Chromient in Infandy in Infcantion工具仍在其中,其构成的工具是其构成的工具。 (Callaway,2018年)。此外,需要特殊的光源来可视化荧光信号,这增加了荧光标记物的应用成本和不便,尤其是在领域条件下。我们开发了一个可视化工具箱,即Vimebox(Visual Maize Editing Toolbox),用于选择玉米(Zea Mays)中的正变换体。在Vimebox系统中,表达Cas9的矢量包括一个基因盒,该盒子包含从组织特异性启动子表达的可见标记物;通过可见标记易于分离的无CAS9核也正在经过基因编辑。vimebox提供了两个优点:(i)它可以增强dsred2的表达,这使得含有cas9的种子在自然光中可见,并且不会影响基因组编辑的效率或植物的发展。(ii)它对不同种子组织有效,例如,使用特异性启动子或启动子优先表达在胚胎或核龙中。此外,Vimebox在不同的其他场景中还具有潜在的应用。

乌干达看到埃博拉疫苗试验的“最快推出”:WHO
通过分析被鉴定为潜在的治疗靶点的每个基因,研究人员进一步探索了其他生物学证据,以加强蛋白质与心力衰竭亚型之间的因果关系。然后,他们在175,000名非裔美国人,西班牙裔和欧洲血统的多项式遗传学数据集中验证了这些发现,并使用替代蛋白质组学平台复制了结果。


看到神经霉素炎谱障碍的差异:将新型策略纳入护理
最近,研究人员开始专注于AQP4-IGG阴性NMOSD。由杰基[Palace]领导的一个牛津集团最近对这种疾病患者进行了36个临床MRI参数的主要成分分析,他们发现此类别中有三个表型亚组:MS样亚组,NMOSD类似于NMOSD的亚组和低脑部病变亚组。MS样亚组具有MRI病变,通常在MS中看到,其代谢组学研究表明,该组中的肌异醇和甲酸水平明显高于NMOSD样子组。因此,这两组与病理的观点可能不同。NMOSD样子组具有与NMOSD诊断一致的特征。低脑病变亚组的特征是三个或更少的脑病变。因此,血浆NMOSD本质上可能是异质的。

i(看到$ 1 NL©lfd,Anj§$ 160A – 07人畜共患病,矢量...
将在地区和州一级建立强大的数据管理系统,以监测和管理气候敏感疾病的物理,财务和流行病学概况。将在州一级建立一个最先进的健康和气候变化中心,该系统将确保实时数据收集,分析和报告,以跟踪气候变化的疾病事件加剧的疾病事件,例如人畜共患病,媒介疾病,热压力,热压力和呼吸疾病等。此数据的集成将为资源分配提供信息,医疗保健

基于NSUC1610的车载步进电机控制
NSUC1610 是通过反电动势的大小来进行堵转检测,在马达相位未通电期间,可以检测到 BEMF 电压。但这 不包括全步进模式,因为两个相位始终通电。以下假设在微步进模式下检测失速,BEMF 电压与电机转速成 正比,这样可以判断电机是否运行。由于只有在一相未通电的情况下才能进行测量,因此对 BEMF 电压的观 察非常有限。对于理想的电机,在没有任何负载和损耗的情况下,转子将随着定子磁场持续旋转,并且在相电 流为零时,可以看到 BEMF 电压的峰值。对于实际电机和外加负载,转子将始终滞后于定子磁场。此负载相关 相位滞后将导致固定测量点处 BEMF 电压的负载相关变化。在零相位滞后的情况下,可以测量 BEMF 电压峰 值,并且只能看到反电势与速度的相关性。在与负载变化的情况下,反电势会产生相位滞后,BEMF 电压将从 峰值将出现偏移,当这个电压大于或者小于一个阈值时,这就标志着检测到失步点,电机运动将停止。BEMF 电压测量仅在零电流阶跃期间启用。在零电流阶跃结束时,采样和测量最后一次 BEMF 电压值。这可确保线 圈电流达到零,且 BEMF 电压实际可见。根据电机参数、速度和阶跃模式,零阶跃可能会变短,并且无法获得 明显的 BEMF 电压。此时则无法检测失速。失速检测仅在匀速运动期间进行,在加速或减速期间,BEMF 电压 可能非常低,则不会启用失速检测。具体电流波形如图 2.5 所示:

通过数字棱镜看到世界:混合现实中传播视频使用的心理含义
数以百万计的人每天将依靠相机和屏幕来向他们展示周围世界。Apple,Meta和其他公司是大量生产的耳机,可以阻止现实世界中的光,而是依靠PassThrough视频作为混合现实的促成技术。本文的11位作者分别花了几个小时的时间在公共场合和私下戴上这些耳机,其目的是记录PassThrough的经验,然后组织和审查以前的研究,这将有助于研究学者,行业领导者和其他组织随着时间的流逝更好地了解心理后果。首先,我们描述了为什么传道将成为媒体景观的重要组成部分。接下来,我们总结了使新的通行头耳机从以前脱颖而出的技术规范,但与人类对参数的愿景(例如视野,失真,延迟和分辨率)相比,但仍然具有较低的责任感。接下来,我们回顾相关的先前心理学研究。我们得出的结论是,传道经验可以激发敬畏并介绍许多应用,但也可能会引起视觉后效应,距离判断的失误,引起模拟器疾病以及干扰社会联系。我们建议游说日常使用这些耳机的公司谨慎和克制,并敦促学者严格,纵向研究这种现象。
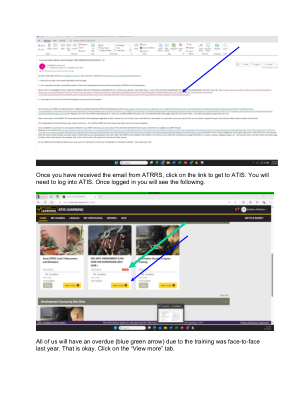
收到 ATRRS 的电子邮件后,点击链接进入 ATIS。您需要登录 ATIS。登录后,您将看到以下内容
收到 ATRRS 的电子邮件后,点击链接进入 ATIS。您需要登录 ATIS。登录后,您将看到以下内容。

在一个仅通过从河流和池塘里抽水而活的水生昆虫动物区系的时代,我们可以看到...
开发了一种用于昆虫的DNA条形码分析的多功能遗传标记,该标记已在全球范围内寻求(我希望将其应用于环境DNA分析,也可以应用于陆生昆虫!)[研究结果]⚫近年来,来自环境DNA的元基因组(社区)分析全面研究了仅通过从河流和池塘中收集水来居住在那里的生物群,在全球范围内吸引了大量关注,需求一直在增加。 ⚫在鱼类,鸟类,哺乳动物和甲壳类动物中已经开发了高度的遗传标记(PCR引物),并经常用于环境DNA分析。 ⚫在昆虫中,地球上种类最多的物种,对全面的社区分析的需求很大,但是物种多样性越高,遗传多样性越高,使得可以开发可用于所有昆虫的基因标记,并且与其他动物群体相比已落后。但是,即使在本文中的试验中,这次发表的论文中发表的遗传标记也已被证明是高度的,而且预计不仅在水生昆虫中,而且昆虫的社区分析都会迅速加速。 ⚫当提交论文时,以预印式的形式发布了有关新开发的标记的信息,并在出版之前已在国内和国际环境DNA分析中使用,并且也因标记的多功能性而受到好评。一些合同进行环境DNA分析工作的公司已经在其网站上发布了日本对本文的解释。 https://edna-blog.com/paper/insectprimer/

“纯粹的快乐”可能不是你期望在目的陈述中看到的第一个短语,但纯粹的快乐是唯一能描述我第一次
“纯粹的喜悦”可能不是你期望在目的陈述中看到的第一个短语,但纯粹的喜悦是描述我第一次改变人类细胞基因组时感受的唯一方式。在我对这些细胞进行测序后,我的分析显示,经过数月的故障排除后,编辑效率仍未达到。这个秘密来自我找到并适应我们系统的新预印本,这意味着我们离理解一种假定的适应性变体在选择下在代谢中的作用如何发挥作用又近了一步。正是这种能够提出以前未知的问题,了解我们周围世界的工作方式,并真正得到答案的能力——即使在多次失败之后——促使我继续我的研究生生涯。除了进化生物学和基因组学之外,我无法想象自己能找到如此有趣的问题来解决,如此激发我整个大脑的问题。杜克大学的遗传学和基因组学系正在提出这些关于现实世界、基础生物学的广泛问题,这一事实让我深感兴奋,能够加入这个研究人员社区,他们不断致力于追求该领域的卓越。我第一次体验到这样一个社区能够理解这种似乎永无止境的求知欲望,那是在我第一次进行实地研究探险的时候。白天,我在落基山脉收集金鱼草杂交花,与维也纳科学技术研究所的 Nick Barton 博士实验室一起进行基因分型。晚上,我在夜间的实地团队晚餐上聆听了几个小时绝对迷人的博士后和研究生们热烈讨论生态学、杂交区和自然选择等各种问题。我只想成为他们中的一员,参与这些对话并做出有意义的贡献。自然而然,这种对科学的热爱让我在两个月后就周末在环境控制室里收集虫卵。从西班牙回来后,我找到了韦尔斯利学院生物系唯一的进化生物学家 Andrea Sequeira 博士。在她的实验室里,我深入研究了一个项目,研究两种克隆繁殖的入侵昆虫物种如何将其基因表达程序适应各种新宿主植物。我们能够观察到基因表达差异与可用宿主植物类型之间的关联,令人惊讶的是,这些基因表达差异在成虫和进食前的后代之间也存在。这是我第一次理解生态学、测序技术和进化生物学如何整合起来,提出任何领域都无法单独解决的问题。我将这个项目从实验台推进到分析阶段,最终完成了我的系荣誉论文、PLOS One 1 上的第一作者出版物,并在 2019 年国际进化会议上介绍了这个项目。在这里,我能够与不同的研究人员进行深入的对话,而这些对话曾经超出了我的理解范围,我们对解读生命复杂性有着共同的兴趣。这让我坚信,研究社区是唯一可以满足我一生继续研究进化问题的愿望的地方。虽然我是在 COVID-19 疫情期间毕业的,但我在麻省理工学院和哈佛大学布罗德研究所的 Pardis Sabeti 博士的实验室里找到了一个可以推动我发挥智力极限的新家。在这里,我开始研究基因组学的一个基本问题:DNA 序列如何影响基因表达?我为我们小组开发高通量 CRISPR 干扰筛选做出了贡献,该筛选可以识别任何基因的非编码调控元件,我作为共同作者在《自然遗传学》杂志上发表了描述该方法的论文 2,这反映了这一点。然后,我开始关注一个相关问题,即这些调控元件内的非编码人类变异如何影响基因表达,并开发了我尖端的分子基因组学方法和计算分析工具。我致力于优化 CRISPR-Cpf1 基因组编辑方法,以测试假定的因果非编码多态性的功能后果。利用这些等位基因