XiaoMi-AI文件搜索系统
World File Search System纤溶

止血手册
ACA抗抑制剂抗体APC激活的蛋白C apla抗磷脂抗体APS抗磷脂抗体综合征在抗凝血酶中; antithrombin III AvWS acquired von Willebrand syndrome CLSI Clinical Laboratory Standards Institute (formerly NCCLS) DIC disseminated intravascular coagulation DOAC direct oral anticoagulant dRVVT dilute Russell viper venom time DTI direct thrombin inhibitor ELISA enzyme-linked immunosorbent assay FDP fibrin degradation products (aka fibrin split products) FEU fibrinogen equivalent units FVL factor V Leiden HIT heparin-induced thrombocytopenia HMWK high-molecular-weight kininogen HMWM high-molecular-weight multimer HSP Henoch-Schönlein purpura INR international normalized ratio ISI international sensitivity index ISTH International Society on Thrombosis and Haemostasis LMWH low molecular weight heparin NHLBI National Heart, Lung, and Blood Institute NIH National Institutes of Health NORD National Organization for Rare Disorders PAI-1 plasminogen activator inhibitor-1 PF4 platelet factor 4 PT prothrombin time PTT partial thromboplastin time RT reptilase time RVVT Russell viper venom time SLE systemic lupus erythematosus SSC Scientific和标准化小组委员会(ISTH的)TEG®血栓射击TFPI组织因子途径抑制剂TPA组织纤溶酶原激活物;组织型纤溶酶原激活剂TT凝血酶时间;凝血酶凝结时间; TCT TCT TTP血栓细胞减少紫菜UFH未分离的肝素VTE静脉血栓栓塞VWD VON WILLEBRAND疾病vwf vwf von von von von von von von willebrand因子

数据表:MCA1396D550产品详细信息
参考文献1。Els Conrath,K。等。(2001)骆驼单域抗体作为双特异性和双价抗体构建体中的模块化建筑单元。J Biol Chem。 276(10):7346-50。 2。 suen,J.L。 等。 (2001)NZB x NZW F 1小鼠中骨髓衍生的树突状细胞对U1a蛋白的自动T细胞反应和U1A蛋白的抗原决定因素的表征。 免疫。 103:301-309。 3。 Hoffmann,S.C。等。 (2007)鉴定Clec12b,骨髓细胞上的抑制性受体。 J Biol Chem。 282(31):22370-5。 4。 Zheng,J。等。 (2007)在Treg抑制作用中免疫的小鼠的血清将DEK鉴定为神经母细胞瘤肿瘤抗原。 BMC免疫。 8:4。 5。 Bahi,A。 &Dreyer,J.L。 (2008)伏伏核中纤溶酶原激活剂的过表达增强了可卡因,苯丙胺和吗啡诱导的奖励和行为敏化。 基因脑行为。 7(2):244-56。 6。 Wrighton,K.H。 等。 (2009)转化生长因子{beta}可以独立于骨形态发生蛋白受体刺激Smad1磷酸化。 J Biol Chem。 284(15):9755-63。 7。 Diefenbacher,M。等。 (2011)DSL1绑扎复合物积极参与可溶性NSF(N-乙基甲米酰胺敏感因子)附着蛋白受体(SNARE)复合物组件在塞雷维氏酵母中的内质网处。 8。J Biol Chem。276(10):7346-50。2。suen,J.L。等。(2001)NZB x NZW F 1小鼠中骨髓衍生的树突状细胞对U1a蛋白的自动T细胞反应和U1A蛋白的抗原决定因素的表征。免疫。103:301-309。 3。 Hoffmann,S.C。等。 (2007)鉴定Clec12b,骨髓细胞上的抑制性受体。 J Biol Chem。 282(31):22370-5。 4。 Zheng,J。等。 (2007)在Treg抑制作用中免疫的小鼠的血清将DEK鉴定为神经母细胞瘤肿瘤抗原。 BMC免疫。 8:4。 5。 Bahi,A。 &Dreyer,J.L。 (2008)伏伏核中纤溶酶原激活剂的过表达增强了可卡因,苯丙胺和吗啡诱导的奖励和行为敏化。 基因脑行为。 7(2):244-56。 6。 Wrighton,K.H。 等。 (2009)转化生长因子{beta}可以独立于骨形态发生蛋白受体刺激Smad1磷酸化。 J Biol Chem。 284(15):9755-63。 7。 Diefenbacher,M。等。 (2011)DSL1绑扎复合物积极参与可溶性NSF(N-乙基甲米酰胺敏感因子)附着蛋白受体(SNARE)复合物组件在塞雷维氏酵母中的内质网处。 8。103:301-309。3。Hoffmann,S.C。等。(2007)鉴定Clec12b,骨髓细胞上的抑制性受体。 J Biol Chem。 282(31):22370-5。 4。 Zheng,J。等。 (2007)在Treg抑制作用中免疫的小鼠的血清将DEK鉴定为神经母细胞瘤肿瘤抗原。 BMC免疫。 8:4。 5。 Bahi,A。 &Dreyer,J.L。 (2008)伏伏核中纤溶酶原激活剂的过表达增强了可卡因,苯丙胺和吗啡诱导的奖励和行为敏化。 基因脑行为。 7(2):244-56。 6。 Wrighton,K.H。 等。 (2009)转化生长因子{beta}可以独立于骨形态发生蛋白受体刺激Smad1磷酸化。 J Biol Chem。 284(15):9755-63。 7。 Diefenbacher,M。等。 (2011)DSL1绑扎复合物积极参与可溶性NSF(N-乙基甲米酰胺敏感因子)附着蛋白受体(SNARE)复合物组件在塞雷维氏酵母中的内质网处。 8。(2007)鉴定Clec12b,骨髓细胞上的抑制性受体。J Biol Chem。 282(31):22370-5。 4。 Zheng,J。等。 (2007)在Treg抑制作用中免疫的小鼠的血清将DEK鉴定为神经母细胞瘤肿瘤抗原。 BMC免疫。 8:4。 5。 Bahi,A。 &Dreyer,J.L。 (2008)伏伏核中纤溶酶原激活剂的过表达增强了可卡因,苯丙胺和吗啡诱导的奖励和行为敏化。 基因脑行为。 7(2):244-56。 6。 Wrighton,K.H。 等。 (2009)转化生长因子{beta}可以独立于骨形态发生蛋白受体刺激Smad1磷酸化。 J Biol Chem。 284(15):9755-63。 7。 Diefenbacher,M。等。 (2011)DSL1绑扎复合物积极参与可溶性NSF(N-乙基甲米酰胺敏感因子)附着蛋白受体(SNARE)复合物组件在塞雷维氏酵母中的内质网处。 8。J Biol Chem。282(31):22370-5。4。Zheng,J。等。 (2007)在Treg抑制作用中免疫的小鼠的血清将DEK鉴定为神经母细胞瘤肿瘤抗原。 BMC免疫。 8:4。 5。 Bahi,A。 &Dreyer,J.L。 (2008)伏伏核中纤溶酶原激活剂的过表达增强了可卡因,苯丙胺和吗啡诱导的奖励和行为敏化。 基因脑行为。 7(2):244-56。 6。 Wrighton,K.H。 等。 (2009)转化生长因子{beta}可以独立于骨形态发生蛋白受体刺激Smad1磷酸化。 J Biol Chem。 284(15):9755-63。 7。 Diefenbacher,M。等。 (2011)DSL1绑扎复合物积极参与可溶性NSF(N-乙基甲米酰胺敏感因子)附着蛋白受体(SNARE)复合物组件在塞雷维氏酵母中的内质网处。 8。Zheng,J。等。(2007)在Treg抑制作用中免疫的小鼠的血清将DEK鉴定为神经母细胞瘤肿瘤抗原。BMC免疫。 8:4。 5。 Bahi,A。 &Dreyer,J.L。 (2008)伏伏核中纤溶酶原激活剂的过表达增强了可卡因,苯丙胺和吗啡诱导的奖励和行为敏化。 基因脑行为。 7(2):244-56。 6。 Wrighton,K.H。 等。 (2009)转化生长因子{beta}可以独立于骨形态发生蛋白受体刺激Smad1磷酸化。 J Biol Chem。 284(15):9755-63。 7。 Diefenbacher,M。等。 (2011)DSL1绑扎复合物积极参与可溶性NSF(N-乙基甲米酰胺敏感因子)附着蛋白受体(SNARE)复合物组件在塞雷维氏酵母中的内质网处。 8。BMC免疫。8:4。5。Bahi,A。&Dreyer,J.L。(2008)伏伏核中纤溶酶原激活剂的过表达增强了可卡因,苯丙胺和吗啡诱导的奖励和行为敏化。基因脑行为。7(2):244-56。6。Wrighton,K.H。 等。 (2009)转化生长因子{beta}可以独立于骨形态发生蛋白受体刺激Smad1磷酸化。 J Biol Chem。 284(15):9755-63。 7。 Diefenbacher,M。等。 (2011)DSL1绑扎复合物积极参与可溶性NSF(N-乙基甲米酰胺敏感因子)附着蛋白受体(SNARE)复合物组件在塞雷维氏酵母中的内质网处。 8。Wrighton,K.H。等。(2009)转化生长因子{beta}可以独立于骨形态发生蛋白受体刺激Smad1磷酸化。J Biol Chem。 284(15):9755-63。 7。 Diefenbacher,M。等。 (2011)DSL1绑扎复合物积极参与可溶性NSF(N-乙基甲米酰胺敏感因子)附着蛋白受体(SNARE)复合物组件在塞雷维氏酵母中的内质网处。 8。J Biol Chem。284(15):9755-63。7。Diefenbacher,M。等。(2011)DSL1绑扎复合物积极参与可溶性NSF(N-乙基甲米酰胺敏感因子)附着蛋白受体(SNARE)复合物组件在塞雷维氏酵母中的内质网处。8。J Biol Chem。 286:25027-38。 Alvarez,M.M。 等。 (2010)人血清中流感A/H1N1/2009抗体的特定识别:一种简单的无病毒ELISA方法。 PLOS ONE。 5:E10176。 9。 Bahi,A。等。 (2008)组织型纤溶酶原激活剂系统在苯丙胺诱导的条件位置偏好灭绝和恢复中的作用。 神经心理药理学。 33:2726-34。 10。 Gunnarsen,K.S。 等。 (2010)可溶性单链T细胞受体的周质表达由伴侣FKPA挽救。 BMC生物技术。 10:8。 11。 Hwang,H.Y。 等。 (2008)高度特异性的抑制C1Q球形与人IgG结合:一种使用工程的单链抗体可变片段来控制和调节经典补体途径的新方法。 mol免疫。 45:2570-80。 12。 de Vooght,L。等。 (2012)功能性的表达和细胞外释放J Biol Chem。286:25027-38。Alvarez,M.M。 等。 (2010)人血清中流感A/H1N1/2009抗体的特定识别:一种简单的无病毒ELISA方法。 PLOS ONE。 5:E10176。 9。 Bahi,A。等。 (2008)组织型纤溶酶原激活剂系统在苯丙胺诱导的条件位置偏好灭绝和恢复中的作用。 神经心理药理学。 33:2726-34。 10。 Gunnarsen,K.S。 等。 (2010)可溶性单链T细胞受体的周质表达由伴侣FKPA挽救。 BMC生物技术。 10:8。 11。 Hwang,H.Y。 等。 (2008)高度特异性的抑制C1Q球形与人IgG结合:一种使用工程的单链抗体可变片段来控制和调节经典补体途径的新方法。 mol免疫。 45:2570-80。 12。 de Vooght,L。等。 (2012)功能性的表达和细胞外释放Alvarez,M.M。等。(2010)人血清中流感A/H1N1/2009抗体的特定识别:一种简单的无病毒ELISA方法。PLOS ONE。 5:E10176。 9。 Bahi,A。等。 (2008)组织型纤溶酶原激活剂系统在苯丙胺诱导的条件位置偏好灭绝和恢复中的作用。 神经心理药理学。 33:2726-34。 10。 Gunnarsen,K.S。 等。 (2010)可溶性单链T细胞受体的周质表达由伴侣FKPA挽救。 BMC生物技术。 10:8。 11。 Hwang,H.Y。 等。 (2008)高度特异性的抑制C1Q球形与人IgG结合:一种使用工程的单链抗体可变片段来控制和调节经典补体途径的新方法。 mol免疫。 45:2570-80。 12。 de Vooght,L。等。 (2012)功能性的表达和细胞外释放PLOS ONE。5:E10176。9。Bahi,A。等。(2008)组织型纤溶酶原激活剂系统在苯丙胺诱导的条件位置偏好灭绝和恢复中的作用。神经心理药理学。33:2726-34。10。Gunnarsen,K.S。 等。 (2010)可溶性单链T细胞受体的周质表达由伴侣FKPA挽救。 BMC生物技术。 10:8。 11。 Hwang,H.Y。 等。 (2008)高度特异性的抑制C1Q球形与人IgG结合:一种使用工程的单链抗体可变片段来控制和调节经典补体途径的新方法。 mol免疫。 45:2570-80。 12。 de Vooght,L。等。 (2012)功能性的表达和细胞外释放Gunnarsen,K.S。等。(2010)可溶性单链T细胞受体的周质表达由伴侣FKPA挽救。BMC生物技术。 10:8。 11。 Hwang,H.Y。 等。 (2008)高度特异性的抑制C1Q球形与人IgG结合:一种使用工程的单链抗体可变片段来控制和调节经典补体途径的新方法。 mol免疫。 45:2570-80。 12。 de Vooght,L。等。 (2012)功能性的表达和细胞外释放BMC生物技术。10:8。11。Hwang,H.Y。 等。 (2008)高度特异性的抑制C1Q球形与人IgG结合:一种使用工程的单链抗体可变片段来控制和调节经典补体途径的新方法。 mol免疫。 45:2570-80。 12。 de Vooght,L。等。 (2012)功能性的表达和细胞外释放Hwang,H.Y。等。(2008)高度特异性的抑制C1Q球形与人IgG结合:一种使用工程的单链抗体可变片段来控制和调节经典补体途径的新方法。mol免疫。45:2570-80。12。de Vooght,L。等。(2012)功能性的表达和细胞外释放

设计和探索您自己的肠溶衣包衣药物
我们的消化系统是我们身体日常功能中最重要的方面之一。我们吃的每一点食物或我们摄入的任何东西都必须分解成可以被身体吸收的营养物质,这就是为什么完全消化食物需要几个小时的原因。在人类中,蛋白质必须分解成氨基酸,淀粉分解成单糖,脂肪分解成脂肪酸和甘油。甚至食物和饮料中的水也会被吸收到血液中,为身体提供所需的液体。这意味着我们的身体会试图分解和吸收所有摄入的东西,即使它不是食物,比如药物。许多药物如果在身体的错误部位吸收或以错误的剂量吸收,可能会很危险。这就是为什么多年来科学家和工程师一直在努力完善药物的生产,以便它们进入消化系统和人体时是安全有效的。

液体混溶间隙对 Inconel 625 缺陷的影响...
摘要:由于铜基合金具有高热导率,而镍基高温合金具有高高温抗拉强度,因此铜基弥散强化合金与镍基高温合金的连接在液体火箭发动机应用中引起了越来越多的关注。然而,这种接头在通过液态过程连接时可能会开裂,从而导致零件失效。在本文中,将 15–95 wt.% GRCop42 成分与 Inconel 625 合金化,并对其进行了表征,以更好地了解开裂的根本原因。结果表明,在对应于 30–95 wt.% GRCop42 的成分中,贫铜液体和富铜液体之间缺乏可混溶性。观察到两种不同的形态,并通过使用 CALPHAD 进行解释; 30–50 wt.% GRCop42 处为铜缺乏的枝晶,枝晶间区域为富铜,60–95 wt.% GRCop42 处为铜缺乏的球体,周围为富铜基质。相分析表明,脆性金属间相在 60–95 wt.% GRCop42 铜缺乏区域析出。本文提出了三种开裂机制,为避免镍基高温合金与铜基弥散强化合金接头缺陷提供指导。

中性粒细胞胞外陷阱和溶栓抵抗
结果:结果显示脑血栓与冠状动脉血栓成分不同,溶解性脑血栓明显多于冠状动脉血栓(66% vs 14%;P =0.005)。脑血栓中NETs的表达明显高于冠状动脉血栓,H2B的表达高于脑血栓(P =0.031)。无论血栓来源如何,血栓溶解均与NE阳性率升高显著相关(平均边际效应为6.461[95%CI,0.7901~12.13];P =0.02555)。调整血栓部位后,抗凝治疗/肝素治疗与H2B/NE量之间无显著相关性。重要的是,血栓年龄是 NET 含量的唯一独立预测因素,无需任何溶栓治疗(P =0.014)。

纳豆激酶封装纳米药物用于靶向溶栓:开发、改善体内溶栓效果以及超声/光声成像
摘要:纳豆激酶 (NK) 是一种强效的溶栓酶,可溶解血栓,在心血管疾病的治疗中被广泛使用。然而,由于其高分子量和蛋白质性质,稳定性和生物利用度问题使其有效输送仍然很困难。在本研究中,我们通过反相蒸发法开发了新型 NK 负载非靶向脂质体 (NK-LS) 和靶向脂质体 (RGD-NK-LS 和 AM-NK-LS)。通过 Zetasizer、SEM、TEM 和 AFM 进行物理化学表征 (粒度、多分散性指数、zeta 电位和形态)。Bradford 测定和 XPS 分析证实了靶向配体的表面结合成功。通过 CLSM、光子成像仪 optima 和流式细胞术进行的血小板相互作用研究表明,靶向脂质体的血小板结合亲和力明显较高 (P < 0.05)。使用人体血液和 CLSM 成像进行的纤维蛋白溶解研究进行了体外评估,证明了 AM-NK-LS 具有强大的抗血栓功效。此外,出血和凝血时间研究表明靶向脂质体没有任何出血并发症。此外,使用多普勒流量计和超声/光声成像对 Sprague-Dawley (SD) 大鼠体内 FeCl 3 模型进行的体内实验表明,靶向脂质体对血栓部位的血栓溶解率增加且具有强大的亲和力。此外,体外血液相容性和组织病理学研究证明了纳米制剂的安全性和生物相容性。关键词:纳豆激酶、血栓溶解、纤维蛋白溶解、血栓靶向、光声成像
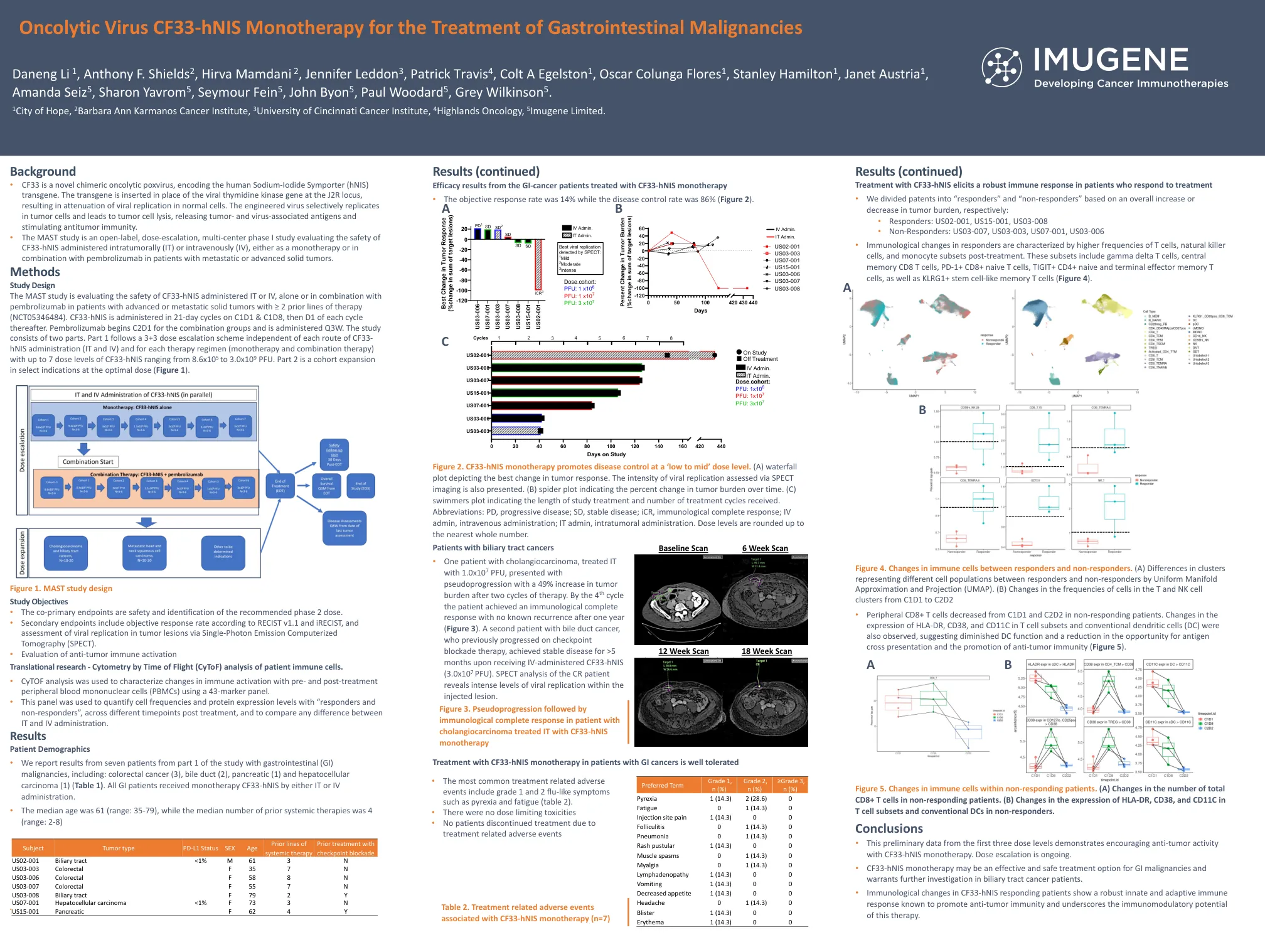
溶瘤病毒CF33-hNIS单一疗法治疗胃肠道恶性肿瘤
背景 • CF33 是一种新型嵌合溶瘤痘病毒,编码人类钠碘转运体 (hNIS) 转基因。转基因插入 J2R 基因座的病毒胸苷激酶基因位置,导致正常细胞中病毒复制减弱。工程病毒选择性地在肿瘤细胞中复制并导致肿瘤细胞裂解,释放肿瘤和病毒相关抗原并刺激抗肿瘤免疫。 • MAST 研究是一项开放标签、剂量递增、多中心 I 期研究,评估 CF33-hNIS 肿瘤内 (IT) 或静脉内 (IV) 给药的安全性,无论是作为单一疗法还是与派姆单抗联合用于转移性或晚期实体瘤患者。方法研究设计 MAST 研究正在评估 CF33-hNIS 单独或与帕博利珠单抗联合通过 IT 或 IV 给药的安全性,适用于既往接受过 ≥ 2 种治疗方案的晚期或转移性实体瘤患者(NCT05346484)。CF33-hNIS 在 C1D1 和 C1D8 以 21 天为一个周期给药,之后每个周期的 D1 给药。联合组的帕博利珠单抗从 C2D1 开始,每三周给药一次。该研究分为两部分。第 1 部分遵循 3+3 剂量递增方案,与每种 CF33- hNIS 给药途径(IT 和 IV)和每种治疗方案(单一疗法和联合疗法)无关,CF33-hNIS 剂量水平最多可达 7 个,范围从 8.6x10 5 至 3.0x10 9 PFU

用于癌症免疫治疗的溶瘤病毒 Hemminki, Otto
在本综述中,我们讨论了溶瘤病毒在癌症免疫疗法中的应用,特别关注腺病毒。这些病毒可作为模型来阐明病毒的多功能性,以及如何将它们用于补充其他癌症疗法以获得最佳患者益处。一百多年的历史报告表明腺病毒和其他溶瘤病毒的治疗效果和安全性。这在更多当代患者系列和多项临床试验中得到了证实。然而,虽然第一批病毒已经获得多个监管机构的批准,但仍有改进的空间。由于已经看到良好的安全性和耐受性,溶瘤病毒领域现已转向提高多种方法的疗效。向病毒中添加不同的免疫调节转基因是一种势头强劲的策略。因此可以在肿瘤处产生免疫刺激分子,同时减少全身副作用。另一方面,临床前研究表明,与放疗和化疗等传统疗法具有附加或协同作用。此外,新推出的检查点抑制剂和其他免疫调节药物可以成为溶瘤病毒的完美伴侣。尤其是那些似乎无法被免疫系统识别的肿瘤,可以通过溶瘤病毒产生免疫原性。从逻辑上讲,与检查点抑制剂的结合正在正在进行的试验中进行评估。另一个有希望的途径是利用溶瘤病毒调节肿瘤微环境,使 T 细胞疗法能够在实体瘤中发挥作用。溶瘤病毒可能是癌症免疫治疗的下一个引人注目的浪潮。

阿托伐他汀的溶出度曲线比较评估...
阿托伐他汀是一种用于治疗高脂血症的一线药物,该专利于2011年用完。目前,在印尼流通的阿托伐他汀片剂有原创药、多个品牌药和仿制药。本研究通过性能测试、重量均匀度、硬度、易碎性、崩解时间、使用紫外可见分光光度计测定含量和溶出度试验等物理质量测试来确定印尼流通的阿托伐他汀制剂的质量。溶出度试验使用 pH 值为 1.2、4.5 和 6.8 的 aquades 介质和缓冲溶液,体积为 900 ml,搅拌速度为 100 rpm,温度为 37°C ± 0.5,测试时间为 45 分钟。对三个样品进行了测试,即原创药、品牌药和仿制药阿托伐他汀片剂样品。三个药片样品均符合所有物理标准,包括重量均匀性、硬度、易碎性和崩解时间。原研药、品牌药和仿制药的阿托伐他汀含量测定结果均符合含量要求,即不低于标签标示量的 90.0%,不超过标签标示量的 110.0%。品牌药阿托伐他汀片的溶出曲线与原研药相似,而仿制药阿托伐他汀片的溶出曲线与原研药不同。关键词:阿托伐他汀,物理质量测试,溶出曲线
心心心肌病:一种疾病J-Wave综合征和特发性心室纤颤
本文中表达的观点不一定是编辑或美国心脏协会的观点。通讯:Elijah R Behr,马萨诸塞州,医学博士,心血管临床学术组,分子与临床科学研究所,圣乔治大学,伦敦圣乔治大学,克兰默露台,伦敦,SW17 0RE,EBEHR@SGHR@SGUL.AC.AC.AC.UK;或Amsterdam UMC的Ruben Coronel,位置AMC,Meibergdreef 9,1105 AZ Amsterdam,荷兰,电子邮件r.coronel@amsterdamumc.nl *r。 Coronel和E.R.behr同等贡献。有关资金和披露的来源,请参见第1630页。©2023作者。流通于美国心脏协会,Inc。发表这是根据Creative Commons归因许可条款的开放访问文章,该条款允许在任何媒介中使用,分发和复制,前提是适当地引用了原始作品。