XiaoMi-AI文件搜索系统
World File Search System细胞分裂

重组 Cenarchaeum symbiosum DNA 聚合酶 II 大亚基 (polC),部分
Cenarchaeum symbiosum DNA 聚合酶 II 大亚基 (polC) 是古菌 Cenarchaeum symbiosum 中 DNA 复制过程中的关键酶。其主要功能包括在细胞分裂和基因组维持过程中合成 DNA 链。其研究领域包括揭示其在基因组稳定性中的作用以及探索其在研究古细菌遗传学中的应用。在分子生物学和古细菌遗传学中,polC 是理解 DNA 复制机制的关键焦点。该酶的重要性在于提供对基本生物过程的洞察,促进古细菌分子生物学的进步。
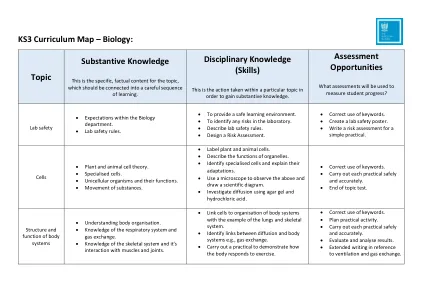
KS3课程图 - 生物学:主题实质知识
•解释如何发生变化。•解释特征是继承,环境还是两者兼而有之。•描述环境变化和继承变化之间的差异。•研究变化和分析数据。•能够使用图形数据来查看连续变化和不连续变化之间的关系。•回忆关键字的定义:染色体,基因和DNA,并链接到其功能。•描述特征是如何继承的。•确定在细胞分裂,性细胞产生和受精过程中染色体的数量如何变化。•描述科学家如何共同开发DNA模型。•解释DNA的变化如何影响生物体及其未来后代。

原住民便利化的典范:开放、赋权……
此外,分组讨论会不会分散到不同的地点。就像细胞分裂一样,小组从主圈开始形成,所有小组的参与者都可以通过聚集在每个节点周围的参与者数量来了解哪些主题最有趣。与更传统的引导技术相反,固执己见或咄咄逼人的参与者通常会主导会议进程,但最终会发现自己没有观众,因为参与者可以自由地转向他们最感兴趣的讨论。这是一个有机的过程,也是高度民主的。即使是胆小或不情愿的参与者也可以发现自己能够直接影响事件,只要参加他们认为有趣或重要的讨论,而不参加那些不有趣的讨论。

植物着丝粒的结构、功能和进化
着丝粒是真核染色体的重要区域,负责形成着丝粒复合体,在细胞分裂过程中与纺锤体微管连接。值得注意的是,尽管着丝粒在染色体分离中保持保守功能,但其底层 DNA 序列在物种内和物种间都存在差异,并且主要是重复性的。着丝粒的重复内容包括高拷贝串联重复序列(卫星)和/或特定的转座子家族。着丝粒的功能区域由特定组蛋白 3 变体 (CENH3) 的加载定义,该变体使着丝粒成核并显示动态调节。在许多植物中,着丝粒由卫星重复阵列组成,这些阵列的 DNA 甲基化程度高,并被嗜着丝粒的逆转录转座子侵入。在某些情况下,逆转录转座子成为 CENH3 加载的位点。我们回顾了植物着丝粒的结构,包括单着丝粒、全着丝粒和元多着丝粒结构,这些结构在染色体上着丝粒附着位点的数量和分布上有所不同。我们讨论了 CENH3 负荷的变化如何在植物胚胎发生早期细胞分裂过程中驱动基因组消除。我们回顾了表观遗传状态如何影响着丝粒身份,并讨论了试图解释跨物种观察到的着丝粒序列快速变化的进化模型,包括重组的潜在作用。我们概述了可能在着丝粒内起作用的假定选择模式,以及重复序列在驱动着丝粒进化周期中的作用。虽然我们的主要重点是植物基因组,但我们将其与动物和真菌着丝粒进行了比较,以得出着丝粒结构和功能的真核生物范围的视角。

TSH和CTBP相互作用坐标果蝇眼发育
保守转录因子的不同组合调节眼睛前体细胞的分裂,然后在果蝇(果蝇)幼虫前体组织中诱导感光细胞规范,称为眼盘。在第三龄幼虫寿命中,由凹入细胞层制成的形态发生沟(MF)起源于眼盘后缘,并朝着眼盘前侧传播。MF前面的细胞处于增殖阶段,其后部细胞开始分化为感光体。分化的视网膜细胞形成果蝇中化合物成年眼睛的单位。先前的研究表明,锌指转录因子(TSH)促进了MF前方的细胞分裂。C末端结合蛋白(CTBP)是一种保守的转录共抑制剂,可限制眼盘中的细胞分裂。有趣的是,我们的免疫沉淀分析表明,TSH和CTBP分子在眼盘中相互作用。因此,我们的研究目标是确定分子相互作用是否与果蝇中的眼睛发育途径相关。我们已经开发了蝇菌株,在MF前部的分裂细胞中TSH&CTBP过表达。结果,我们发现苍蝇中没有TSH过度表达的苍蝇中没有或微小的成年眼睛,并且在CTBP过表达的苍蝇中出现了微妙的较大的成年眼。接下来,我们计划通过过度表达TSH&CTBP来评估其相互作用对眼表型的影响来制作双突变体。结果将有助于确定由TSH和CTBP调节的眼睛发育过程。

COVID-19 疫苗和输血
英国批准使用的疫苗符合 MHRA 制定的严格安全、质量和有效性标准。任何获批的 COVID-19 疫苗都必须经过所有其他获批药物都要经过的所有临床试验和安全检查。COVID-19 疫苗含有 mRNA 或经过修饰的腺病毒 DNA,它们不会复制,也不会在细胞分裂时被复制。虽然少量疫苗可能会进入血液,但这对疫苗的作用并不重要,而且任何确实发生的血液渗透都可能是短暂的。英国批准的 COVID-19 疫苗不会对血液供应的安全或接受输血的患者构成任何风险。
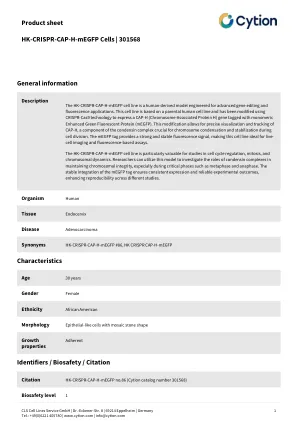
产品表Cellule U2OS-CRISPR-NUP96-HALO | 300448产品表Komórkiu2os-crispr-nup96-snap产品表célulasasb-xiv | 400120产品表HK-Crispr-CAP-H-MEGFP细胞| 301568
HK-CRISPR-CAP-H-MEGFP细胞系是一种为晚期基因编辑和荧光应用而设计的人类衍生模型。该细胞系基于亲本人类细胞系,并已使用CRISPR-CAS9技术进行了修改,以表达用单体增强的绿色荧光蛋白(MEGFP)标记的CAP-H(染色体相关蛋白H)基因。这种修饰允许CAP-H的精确可视化和跟踪,CAP-H是冷凝蛋白复合物的组成部分,对于细胞分裂期间的染色体凝结和稳定至关重要。MEGFP标签提供了强烈而稳定的荧光信号,这使该细胞系非常适合活细胞成像和基于荧光的测定。


植物着丝粒的结构、功能和进化
着丝粒是真核染色体的重要区域,负责形成着丝粒复合体,在细胞分裂过程中与纺锤体微管连接。值得注意的是,尽管着丝粒在染色体分离中保持保守功能,但其底层 DNA 序列在物种内和物种间都存在差异,并且主要是重复性的。着丝粒的重复内容包括高拷贝串联重复序列(卫星)和/或特定的转座子家族。着丝粒的功能区域由特定组蛋白 3 变体 (CENH3) 的加载定义,该变体使着丝粒成核并显示动态调节。在许多植物中,着丝粒由卫星重复阵列组成,这些阵列的 DNA 甲基化程度高,并被嗜着丝粒的逆转录转座子侵入。在某些情况下,逆转录转座子成为 CENH3 加载的位点。我们回顾了植物着丝粒的结构,包括单着丝粒、全着丝粒和元多着丝粒结构,这些结构在染色体上着丝粒附着位点的数量和分布上有所不同。我们讨论了 CENH3 负荷的变化如何在植物胚胎发生早期细胞分裂过程中驱动基因组消除。我们回顾了表观遗传状态如何影响着丝粒身份,并讨论了试图解释跨物种观察到的着丝粒序列快速变化的进化模型,包括重组的潜在作用。我们概述了可能在着丝粒内起作用的假定选择模式,以及重复序列在驱动着丝粒进化周期中的作用。虽然我们的主要重点是植物基因组,但我们将其与动物和真菌着丝粒进行了比较,以得出着丝粒结构和功能的真核生物范围的视角。

胰岛素抵抗和阿尔茨海默氏病
在1916年发现胰岛素受体(IRS)和随后胰岛素降血糖作用的证明中,IRS在控制外周组织中控制葡萄糖代谢方面的关键功能[1,2]。 通过将包括GLUT 4在内的葡萄糖转运蛋白的易位升至质膜,胰岛素可以增强葡萄糖转运到细胞中,并促进外周组织中的葡萄糖利用率。 除了葡萄糖代谢外,胰岛素还会影响蛋白质的合成,细胞分裂和生长。 从历史上看,人们认为大脑是胰岛素不敏感的器官,IR功能主要是外围的。 从观察到循环胰岛素水平似乎对全脑葡萄糖的吸收没有影响的观察得出[3]。 但是,最后一次进行的研究在1916年发现胰岛素受体(IRS)和随后胰岛素降血糖作用的证明中,IRS在控制外周组织中控制葡萄糖代谢方面的关键功能[1,2]。通过将包括GLUT 4在内的葡萄糖转运蛋白的易位升至质膜,胰岛素可以增强葡萄糖转运到细胞中,并促进外周组织中的葡萄糖利用率。除了葡萄糖代谢外,胰岛素还会影响蛋白质的合成,细胞分裂和生长。从历史上看,人们认为大脑是胰岛素不敏感的器官,IR功能主要是外围的。从观察到循环胰岛素水平似乎对全脑葡萄糖的吸收没有影响的观察得出[3]。但是,最后一次进行的研究