XiaoMi-AI文件搜索系统
World File Search System肥鱼

斑马鱼相对Danio Albolineatus
抽象的动物色素模式在行为中起着重要作用,在许多物种中,红色是伴侣选择中个体质量的诚实信号。在Danio鱼类中,有些物种会形成红细胞,含有红色酮心反非的色素细胞,而其他物种,例如斑马鱼(D。Rerio),只有黄色的黄蜂。在这里,我们使用Pearl Danio(D。albolineatus)评估了红细胞的发育起源及其差异机制。我们表明,白化芽孢杆菌的鳍中的红细胞与黄蜂共有一个共同的祖细胞,即使分化后,也可以在细胞命运中保持可塑性。我们进一步确定了将红色颜色赋予红色色彩的主要酮类苯酚,并使用反向遗传学来确定这些细胞分化和维持所需的基因。我们的分析是定义Danio中红色介导的红色色彩发展的机制的第一步,并揭示了与鸟类红色机制的惊人相似之处。

实验之前斑马鱼的适应
根据欧洲议会的指令(2010/63/eu)和针对科学目的的动物保护理事会的指令,附件III点3.1.a,新动物的引入应成为每个设施必须拥有的健康战略的一部分。在第3.7点,还要求设施必须为动物制定习惯计划。此外,委员会的建议2007年6月18日关于用于实验和其他科学目的的动物的适应和护理指南(2007/526/ec),一般部分,4.4指出,允许动物允许动物从运输压力中恢复到新的环境,并习惯新的环境,并习惯了新的环境和新的丈夫和护理习惯。根据委员会的建议,即使被认为身体健康,也必须进行适应时期。所需的时间量取决于动物所经历的东西。例如,与该国境内短期运输相比,长期的国际运输破坏了动物的昼夜节律,可能需要更长的适应期。

深入研究鱼过敏原免疫疗法
鱼过敏是全世界食物过敏原的“九大”类别之一,随着对这种营养食品来源的需求,其流行率正在增加。鱼过敏是一个重大的健康问题,因为它是食品过敏反应的主要原因,占过敏反应死亡的9%。目前治疗鱼过敏的差距是对鱼过敏原的不完全鉴定,在临床环境中缺乏对鱼类过敏原的成分分辨诊断,以及基于不同鱼类消耗实践的敏化分布的可变性。过敏原免疫疗法(AIT)提高了意外食用鱼类的耐受性,并且比药物疗法更长。当前对鱼类AIT的实践或研究范围从口服脱敏到使用纯化的重组白细胞蛋白及其低过敏性变体,被动IgG免疫,并通过改变养殖鱼类饮食的饮食来修饰白蛋白的过敏性。但是,在AIT背景下,基于鱼类过敏原的研究的重点仅限于白蛋白蛋白。需要进行更多的研究才能了解其他鱼类过敏原的参与以及其他几种AIT策略,包括肽疫苗,DNA疫苗,杂交过敏原以及使用具有多种过敏原的纳米体的使用。对于AIT,要考虑的其他重要方面是脱敏的途径,以及评估免疫疗法成功的生物标志物。最后,我们还解决了FISH AIT的几个临床注意事项。

组蛋白乙酰化位点在心脏肥大和心力衰竭中的作用
心力衰竭是由导致心脏肥大的各种生理和病理刺激引起的。这种病理过程常见于多种心血管疾病,并最终导致心力衰竭。心脏肥大和心力衰竭的发展涉及基因表达的重编程,这一过程高度依赖于表观遗传调控。组蛋白乙酰化受心脏应激的动态调节。组蛋白乙酰转移酶在心脏肥大和心力衰竭的表观遗传重塑中起重要作用。组蛋白乙酰转移酶的调控是信号转导和下游基因重编程之间的桥梁。研究心脏肥大和心力衰竭中组蛋白乙酰转移酶和组蛋白修饰位点的变化将为治疗这些疾病提供新的治疗策略。本综述总结了组蛋白乙酰化位点和组蛋白乙酰化酶与心脏肥大和心力衰竭的关联,重点介绍了组蛋白乙酰化位点。
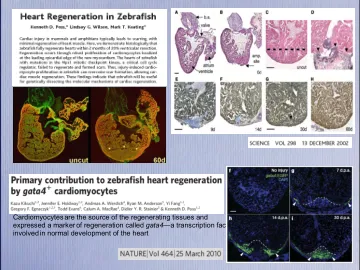
斑马鱼中的心脏再生是
•首先,可能会刺激现有肌纤维中的差异化CMS,以进入细胞周期,分裂和改革顶点。•第二,可以通过募集形成新的增生性CM的未分化的祖细胞来进行再生。•关于再生肌肉起源的第三个可能的机制是这两种称为“去分化”的机制的嵌合体,其中现有肌肉将下调收缩基因以创建未分化或不良分化的细胞。

斑马鱼神经素项目数据库
描述:成人斑马鱼模型的神经行为和生理数据的数据库,通过为斑马鱼遗传信息提供了可用的存储库,通过提供动态的,开放的访问数据存储库,这些数据库是全面的,经过精心策划的Zebrafish Neurobafish Neurobobehavioral实验的结果收集的。截至2012年5月,它包含超过4500多个实验结果,来自75多种独特的生理和行为测试以及330种不同的药物治疗。ZNP结合了该领域发表的工作的经过验证和策划的数据,以提高对使用成人斑马鱼模型有兴趣的研究人员的当前知识的可访问性。总体而言,该计划将允许研究人员快速审查数据,并使用这些模型指导他们的研究。数据和协议提交现在正在接受。

和 keap1b 基因敲除斑马鱼
Keap1 – Nrf2 通路是一种进化保守的机制,可保护细胞免受氧化应激和亲电试剂的侵害。在稳态条件下,Keap1 与 Nrf2 相互作用并导致其快速蛋白酶体降解,但当细胞暴露于氧化应激/亲电试剂时,Keap1 会感知它们,导致 Keap1 – Nrf2 相互作用不当和 Nrf2 稳定。因此,Keap1 被认为是 Nrf2 激活的“抑制剂”和“应激传感器”。有趣的是,鱼类和两栖动物有两种 Keap1(Keap1a 和 Keap1b),而哺乳动物、鸟类和爬行动物只有一种。系统发育分析表明,哺乳动物 Keap1 是鱼类 Keap1b 的直系同源物,而不是 Keap1a。在本研究中,我们使用斑马鱼遗传学研究了 Keap1a 和 Keap1b 之间的差异和相似之处。我们构建了 keap1a 和 keap1b 的斑马鱼基因敲除系。两种基因敲除系的纯合突变体均可存活且可育。在两种突变幼虫中,Nrf2 靶基因的基础表达和抗氧化活性均以 Nrf2 依赖的方式上调,表明 Keap1a 和 Keap1b 均可作为 Nrf2 抑制剂发挥作用。我们还分析了 Nrf2 激活剂萝卜硫素对这些突变体的影响,发现 keap1a- ,而非 keap1b- ,基因敲除幼虫对萝卜硫素有反应,表明两种 Keap1 的压力/化学感应能力不同。


预测鱼对的长期集体行为
现代计算增强了我们对社会相互作用如何塑造动物社会中集体行为的理解。尽管分析模型在研究集体行为方面占主导地位,但本研究介绍了一个深度学习模型,以评估鱼类杜鹃花的社交相互作用。我们将深度学习方法的结果与实验以及最先进的分析模型的结果进行了比较。为此,我们提出了一种系统的方法来评估集体运动模型的信仰,利用了一组严格的个人和集体时空可观察物。我们证明,社交互动的机器学习模型可以直接与他们的分析同行竞争,以复制微妙的实验可观察物。更重要的是,这项工作强调了在不同时间尺度上进行一致验证的必要性,并确定了关键的设计方面,使我们能够捕捉短期和长期动态的深度学习方法。我们还表明,我们的方法可以扩展到没有任何培训的情况下以及其他鱼类,同时保留了深度学习网络的相同结构。最后,我们讨论了在动物群体中集体运动研究的背景下,ML的附加值及其作为分析模型的补充方法的潜力。

鱼免疫能力足以面对气候变化?
持续的气候变化已经与野生鱼类和养殖鱼类的疾病爆发增加有关。在这里,我们评估了当前关于气候变化相关的生态免疫学的知识,重点是探索多种压力源的交互作用,重点是临时,缺氧,盐度和酸化。我们的文献综述表明,温度和溶解氧的急性和慢性变化会损害鱼类免疫力,从而导致疾病易感性增加。此外,已经证明温度和缺氧可以增强某些病原体/寄生虫的感染并加速疾病进展。也很少有针对酸化的研究,但是直接的免疫作用似乎受到限制,而盐度研究导致了对比结果。同样,对于揭示同时改变环境因素的相互作用所必需的多压力实验仍然很少。这最终阻碍了我们估计气候变化在多大程度上会妨碍鱼类免疫力的能力。我们对表观遗传调节机制的评论突出了鱼类免疫反应对不断变化的环境的适应潜力。但是,由于表观遗传学研究数量有限,因此无法得出总体结论。最后,我们提供了如何更好地估计鱼类未来免疫研究的现实气候变化情景影响的前景。