XiaoMi-AI文件搜索系统
World File Search System腺样体

2019财年专利申请技术趋势调查(航天器)
中国空间技术研究院 (中国) 643 26,135 30 空客 (欧洲) 611 13,954 67 波音 (美国) 430 14,624 88 Energiya (俄罗斯) 430 7,401 37 三菱电机 279 89,137 20 IHI 201 13,657 28 泰雷兹 (欧洲) 153 6,495 54 三菱重工 131 27,823 16 霍尼韦尔 (美国) 117 19,431 7 雷神 (美国) 105 5,383 3 斯奈克玛 (欧洲) 102 4,363 6 太空系统/劳拉 (美国) 58 168 12 Viasat (美国) 1 685 0 蓝色起源 (美国) 12 19 1 SpaceX(美国) 1 10 9 Rocket Lab(美国) 5 5 0 北京零度空间科技公司(中国) 2 24 0 Mojave Aerospace Ventures(美国) 2 2 0 PLD space(西班牙) 0 0 0 Reaction Engines(英国) 6 13 4 Relativity Space(美国) 0 2 0 Skyrora(英国) 0 0 0 Oneweb(美国) 11 29 0 Blacksky(美国) 0 0 0 Capella Space(美国) 0 0 0 Hawkeye360(美国) 0 6 0 Iceye(芬兰) 0 1 0 OHB System(德国) 1 8 20 Planet(美国) 5 27 2 Spire Global(美国) 6 22 0 ispace(日本) 7 13 1 Planetary Resources(美国) 4 4 1 Astroscale 12 12 0 D-Orbit (意大利) 4 4 0 NASA (美国) 91 1,924 959 日本宇宙航空研究开发机构 119 500 473 国防科技大学 (中国) 69 6,274 280 哈尔滨工业大学 (中国) 338 25,237 274 加州理工学院 (美国) 19 2,648 314 韩国航空宇宙研究院 (韩国) 436 2,739 72

核融合反应堆高温超导导体开发的现状
名启博:プラマ・核融合学志92,396(2016)。[4 W.H.fietz and al。,IEEE Trans。苹果。超级。26,4800705(2016)。 [5]P。Bruzzone和Al。 ,ncle。 Fuance 58,103001(2018)。 l。米切尔和阿尔。 ,超级条件。 SCI。 树。 34,103001(2021)。 !t。安多和al。 ,技术完整。 1,791(1998)。 Lage F. Dahlgren和Al。 ,Eng已满。 甲板。 167,139(2006)。 ]H。H. Hashizume和Al。 ,Eng已满。 甲板。 63,449(2002)。 [10! Y. Ogawa和Al。 ,J。 填充完整的等离子体。 79,643(2003)。 <+11 Z. Yoshida和Al。 ,Ressing主题等离子体。 1,8(2006)。 [12 Y. Ogawa和Al。 ,Ressing主题等离子体。 9,140,014(2014)。 13 V. Corat和Al。 ,Eng已满。 甲板。 136,1597(2018)。 14 A. Sagara和Al。 ,Eng已满。 甲板。 89,2114(2014)。 15 Y. Zhai和Al。 ,Eng已满。 甲板。 135,324(2018)。 https://typeoneergy.com/ [20! Sorbon和Al。 ,Eng已满。 甲板。 100,378(2015)。 [22 A A. Sykes和Al。26,4800705(2016)。[5]P。Bruzzone和Al。,ncle。Fuance 58,103001(2018)。l。米切尔和阿尔。,超级条件。SCI。 树。 34,103001(2021)。 !t。安多和al。 ,技术完整。 1,791(1998)。 Lage F. Dahlgren和Al。 ,Eng已满。 甲板。 167,139(2006)。 ]H。H. Hashizume和Al。 ,Eng已满。 甲板。 63,449(2002)。 [10! Y. Ogawa和Al。 ,J。 填充完整的等离子体。 79,643(2003)。 <+11 Z. Yoshida和Al。 ,Ressing主题等离子体。 1,8(2006)。 [12 Y. Ogawa和Al。 ,Ressing主题等离子体。 9,140,014(2014)。 13 V. Corat和Al。 ,Eng已满。 甲板。 136,1597(2018)。 14 A. Sagara和Al。 ,Eng已满。 甲板。 89,2114(2014)。 15 Y. Zhai和Al。 ,Eng已满。 甲板。 135,324(2018)。 https://typeoneergy.com/ [20! Sorbon和Al。 ,Eng已满。 甲板。 100,378(2015)。 [22 A A. Sykes和Al。SCI。树。 34,103001(2021)。 !t。安多和al。 ,技术完整。 1,791(1998)。 Lage F. Dahlgren和Al。 ,Eng已满。 甲板。 167,139(2006)。 ]H。H. Hashizume和Al。 ,Eng已满。 甲板。 63,449(2002)。 [10! Y. Ogawa和Al。 ,J。 填充完整的等离子体。 79,643(2003)。 <+11 Z. Yoshida和Al。 ,Ressing主题等离子体。 1,8(2006)。 [12 Y. Ogawa和Al。 ,Ressing主题等离子体。 9,140,014(2014)。 13 V. Corat和Al。 ,Eng已满。 甲板。 136,1597(2018)。 14 A. Sagara和Al。 ,Eng已满。 甲板。 89,2114(2014)。 15 Y. Zhai和Al。 ,Eng已满。 甲板。 135,324(2018)。 https://typeoneergy.com/ [20! Sorbon和Al。 ,Eng已满。 甲板。 100,378(2015)。 [22 A A. Sykes和Al。树。34,103001(2021)。!t。安多和al。,技术完整。1,791(1998)。Lage F. Dahlgren和Al。,Eng已满。甲板。167,139(2006)。]H。H. Hashizume和Al。,Eng已满。甲板。63,449(2002)。[10! Y. Ogawa和Al。,J。填充完整的等离子体。79,643(2003)。<+11 Z. Yoshida和Al。,Ressing主题等离子体。1,8(2006)。[12 Y. Ogawa和Al。,Ressing主题等离子体。9,140,014(2014)。13 V. Corat和Al。,Eng已满。甲板。136,1597(2018)。14 A. Sagara和Al。 ,Eng已满。 甲板。 89,2114(2014)。 15 Y. Zhai和Al。 ,Eng已满。 甲板。 135,324(2018)。 https://typeoneergy.com/ [20! Sorbon和Al。 ,Eng已满。 甲板。 100,378(2015)。 [22 A A. Sykes和Al。14 A. Sagara和Al。,Eng已满。甲板。89,2114(2014)。 15 Y. Zhai和Al。 ,Eng已满。 甲板。 135,324(2018)。 https://typeoneergy.com/ [20! Sorbon和Al。 ,Eng已满。 甲板。 100,378(2015)。 [22 A A. Sykes和Al。89,2114(2014)。15 Y. Zhai和Al。 ,Eng已满。 甲板。 135,324(2018)。 https://typeoneergy.com/ [20! Sorbon和Al。 ,Eng已满。 甲板。 100,378(2015)。 [22 A A. Sykes和Al。15 Y. Zhai和Al。,Eng已满。甲板。135,324(2018)。https://typeoneergy.com/ [20!Sorbon和Al。,Eng已满。甲板。100,378(2015)。[22 A A. Sykes和Al。,ncle。Fusion 58,016039(2018)。<3- y。歌曲和Al。 ,Eng已满。 甲板。 183,113247(2022)。 24-24 N. Yanagi和Al。 ,Ressing主题等离子体。 9,140,013(2014)。 ,Proc。 14th Symp。 Fusion Technology,1727(1986)。歌曲和Al。,Eng已满。甲板。183,113247(2022)。24-24 N. Yanagi和Al。 ,Ressing主题等离子体。 9,140,013(2014)。 ,Proc。 14th Symp。 Fusion Technology,1727(1986)。24-24 N. Yanagi和Al。,Ressing主题等离子体。9,140,013(2014)。,Proc。 14th Symp。 Fusion Technology,1727(1986)。,Proc。14th Symp。Fusion Technology,1727(1986)。

FDA/CBER 组织和先进疗法办公室 (...
– 离体转基因细胞 – 非病毒载体(例如质粒) – 复制缺陷型病毒载体(例如腺病毒、腺相关病毒、慢病毒) – 复制能力强的病毒载体(例如麻疹、腺病毒、牛痘) – 微生物载体(例如李斯特菌、沙门氏菌) 干细胞/干细胞衍生

一种可解释的 CNN-Transformer,用于使用 ECG 诊断儿童睡眠呼吸暂停
摘要 — 儿童阻塞性睡眠呼吸暂停 (OSA) 是一种常见的呼吸系统疾病,与心血管风险增加有关。标准诊断是多导睡眠图,但其复杂性、成本和不便性导致诊断不足。为了解决这种情况,我们首次提出了一种简化的替代方法,使用夜间心电图 (ECG) 和基于卷积神经网络和变压器网络的混合模型来估计儿童 OSA 的严重程度。此外,提出了梯度加权类激活映射 (GradCAM) 方法来解释模型结果。为了开展这项研究,使用了来自儿童腺样体扁桃体切除术试验 (CHAT) 和芝加哥大学 (UofC) 数据库的 2,591 条记录。该模型在 CHAT 中实现了 4 类 Cohen's Kappa 0.392,在 UofC 中实现了 0.346。 GradCAM 突出显示了心动过缓-心动过速模式、PQ 和 QT 段以及已识别的 U 波。因此,这种方法可以改善儿童 OSA 的诊断并提供新的相关心脏信息,从而鼓励在临床环境中采用自动化系统。
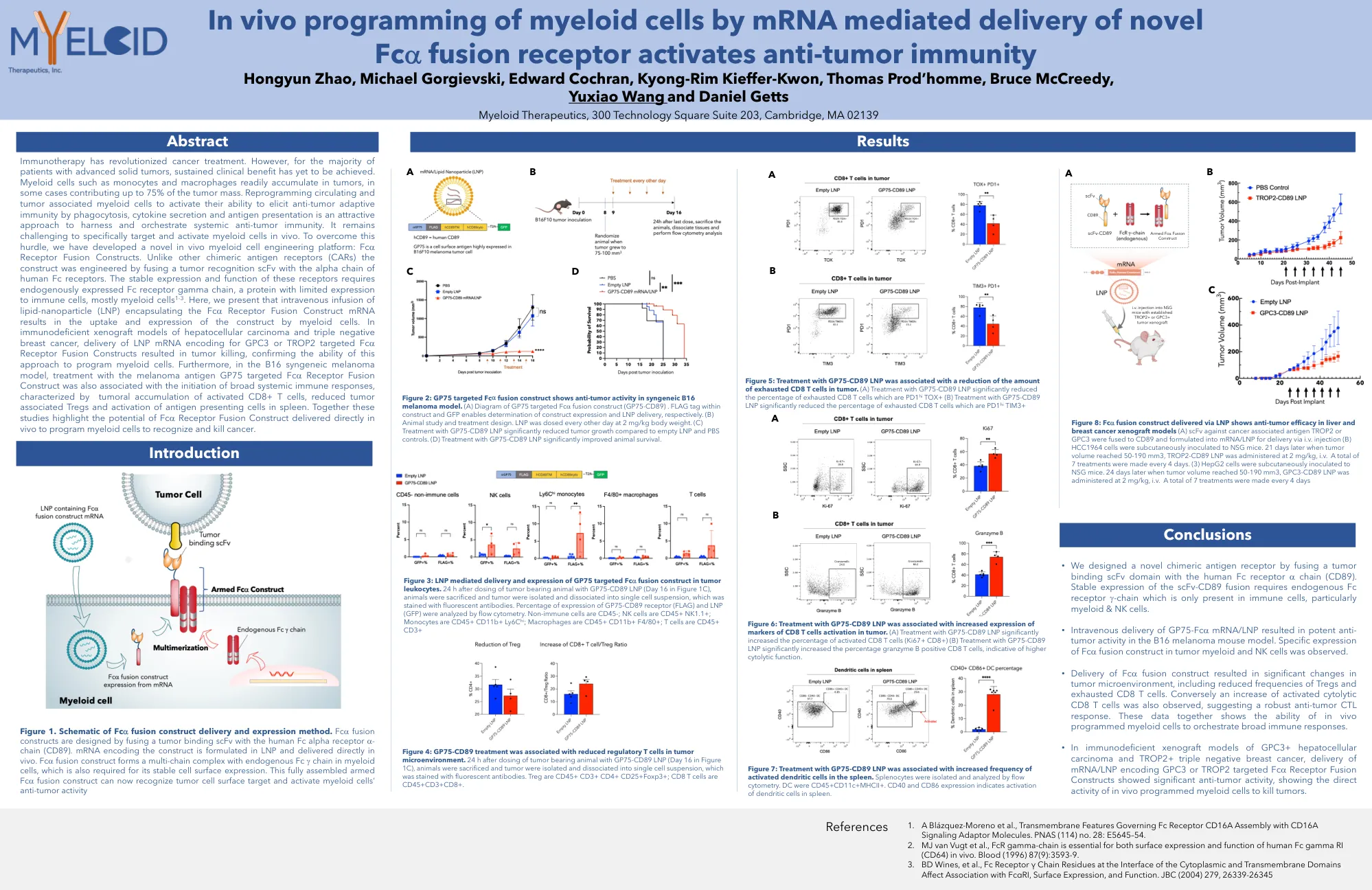
AACR2022体内递送海报
免疫疗法彻底改变了癌症治疗。但是,对于大多数晚期实体瘤患者,尚未实现持续的临床益处。髓样细胞(如单核细胞和巨噬细胞)很容易积聚在肿瘤中,在某些情况下,肿瘤质量的75%。重编程循环和肿瘤与髓样细胞相关,以激活其通过吞噬作用,细胞因子分泌和抗原表现来激活抗肿瘤适应性免疫的能力,是一种有吸引力的方法,可利用并策划系统性的抗肿瘤免疫。在体内专门靶向和激活髓样细胞仍然具有挑战性。为了克服这一障碍,我们开发了一种新型的体内髓细胞工程平台:FC A受体融合构建体。与其他嵌合抗原受体(CAR)不同,该构建体是通过将肿瘤识别SCFV与人体FC受体的α链融合而设计的。这些受体的稳定表达和功能需要内源表达的FC受体γ链,FC受体γ链是一种对免疫细胞表达有限的蛋白质,主要是髓样细胞1-3。在这里,我们介绍了包裹FC A受体融合构建体mRNA的静脉输注脂质 - 纳米颗粒(LNP)导致髓样细胞对构建体的摄取和表达。在肝细胞癌和三重阴性乳腺癌的免疫缺陷异种移植模型中,针对GPC3或trop2靶向FC的LNP mRNA的递送A受体融合构建体导致肿瘤杀死,从而确认了这种方法为骨髓细胞编程的能力。此外,在B16合成性黑色素瘤模型中,用黑色素瘤抗原GP75靶向FC A受体融合构建体的治疗也与启动广泛的全身免疫反应的启动有关,其特征在于肿瘤积累活化的CD8+ T细胞,可减少与肿瘤相关的TREG和SpleeNing spleen and spleen spleen and spleen的活化。这些研究共同强调了FC A受体融合构建体的潜力,直接在体内传递以编程髓样细胞以识别和杀死癌症。

针对靶向药物的工程外泌体
1981年,Trams等。 通过透射电子显微镜发现了一组直径为40-1000 nm的囊泡样结构[1]。 后来,Johnstone等。 在网状细胞成熟过程中鉴定出类似囊泡样的结构,并通过以100,000×g的超速离心为90分钟将这些膜结合的囊泡从绵羊网状细胞中分离出来。 首次将这些囊泡样结构命名为外泌体[2,3]。 但是,当时,外泌体的发现并没有得到太多的关注,因为这些囊泡被认为仅仅是从成熟的红细胞中浪费的产物。 这些囊泡直到最近才被表征为膜结合的细胞外囊泡,在细胞膜与细胞内多囊体(MVBS)融合后通过胞吞作用释放出来[4,5]。 外泌体现在在所有体液和组织中都广泛发现,包括血液[6],尿液[7],母乳[8],羊膜/滑膜/腹水液[9],唾液[10]和脂肪组织[11]。 越来越多的类型的1981年,Trams等。通过透射电子显微镜发现了一组直径为40-1000 nm的囊泡样结构[1]。后来,Johnstone等。在网状细胞成熟过程中鉴定出类似囊泡样的结构,并通过以100,000×g的超速离心为90分钟将这些膜结合的囊泡从绵羊网状细胞中分离出来。首次将这些囊泡样结构命名为外泌体[2,3]。但是,当时,外泌体的发现并没有得到太多的关注,因为这些囊泡被认为仅仅是从成熟的红细胞中浪费的产物。这些囊泡直到最近才被表征为膜结合的细胞外囊泡,在细胞膜与细胞内多囊体(MVBS)融合后通过胞吞作用释放出来[4,5]。外泌体现在在所有体液和组织中都广泛发现,包括血液[6],尿液[7],母乳[8],羊膜/滑膜/腹水液[9],唾液[10]和脂肪组织[11]。越来越多的类型的

AAV 的候选对象和挑战
缩写:AADC,芳香族 L-氨基酸脱羧酶;AAV,腺相关病毒;ALS,肌萎缩侧索硬化症;APOE,载脂蛋白 E;ASO,反义寡核苷酸;ATXN2,共济失调蛋白 2;BBB,血脑屏障;BSCB,血脊髓屏障;CDKL5,细胞周期蛋白依赖性激酶样 5;CNS,中枢神经系统;CRISPR,成簇的规律间隔的短回文重复序列;CSF,脑脊液;DRPLA,齿状红核苍白球路易体萎缩;FTD,额颞痴呆;FUS,聚焦超声;FXTAS,脆性 X 相关震颤/共济失调综合征;GABA,γ-氨基丁酸;GAD,谷氨酸脱羧酶;GAG,糖胺聚糖; GAN,巨轴突性神经病;GBA,葡萄糖脑苷脂酶;GCH,三磷酸鸟苷环化水解酶;GDNF,胶质细胞源性神经营养因子;ICis,脑池内;ICV,脑室内;IPa,脑实质内;IT,鞘内(腰椎);IV,静脉内;LacNAc,硫酸化N-乙酰乳糖胺;MAO,单胺氧化酶;miRNA,微小RNA;MLD,异染性脑白质营养不良;MPS,粘多糖贮积症;MRgFUS,磁共振成像引导聚焦超声;MRI,磁共振成像;MSA,多系统萎缩;NCL,神经元蜡样脂褐素沉积症;NGF,神经生长因子;NTN,神经营养素;PDHD,丙酮酸脱氢酶缺乏症;Put,壳核; rAAV,重组腺相关病毒;RNAi,RNA 干扰;siRNA,短干扰 RNA,小干扰 RNA;SMA,脊髓性肌萎缩;SMARD,脊髓性肌萎缩伴呼吸窘迫;SNc,黑质致密部;SOD1,超氧化物歧化酶 1;Str,纹状体;TDP-43,TAR DNA 结合蛋白 43;TERT,端粒酶逆转录酶;TH,酪氨酸羟化酶;Th,丘脑;VTA,腹侧被盖区;ZFN,锌指核酸酶。 * 通讯作者:德克萨斯大学达拉斯分校,800 West Campbell Road, EW31, Richardson, TX 75080, USA。电子邮箱地址:Zhenpeng.Qin@utdallas.edu (Z. Qin)。

使用全原子模拟和人工智能发现新的淀粉样蛋白样肽
微生物组革命移动了微生物学家的守门柱。几个世纪以来,微生物学一直在理解相对少量的微生物上。这些模型物种是因为它们对健康,环境,工业的重要性,或仅仅是因为该物种易于使用。微生物学家在整个分子,遗传和基因组旋转中保持了关注,但是宏基因组革命使得不可能忽略我们世界各个方面发现的成千上万种研究的物种(DeWhirst等人。2010; Quast等。2013; Parks等。2018)。微生物组的科学崛起令人兴奋,但它给微生物学带来了巨大的实践挑战。如果只花了几个世纪的时间才能学习几种模型物种的细节,我们如何才能理解成千上万的新发现物种?为了说明研究研究的数据的匮乏,我们进行了文献计量分析,以提出微生物学研究的不均匀分布。GTDB数据库的版本202(Parks等人2022)包括43,409种独特的物种,我们计算了参考标题或摘要中每个物种的PubMed文章数量。结果严重偏斜。几乎74%的已知物种从来都不是科学出版物的主题 - 这些是未研究的细菌(图1A)。即使在研究的物种中(至少有一个出版物),所有文章中的50%仅指十种物种(图1b)。因此,我们的知识密度(我们每个物种所学的数量)实际上正在减少。所有细菌学文章中有90%以上研究的物种的研究不足1%,从而产生了细小的微生物的“长尾巴”。科学企业正在扩大,每年科学家发表的论文比久违的年份(国家科学基金会和国家科学委员会2021年)多4-5%。很容易想到,科学产量的增加将克服微生物的长尾巴,也就是说,科学家最终将四处研究每个物种。不幸的是,每年发现的物种数量超过了科学产出的增加(图1C)。在1990 - 2020年之间,每个研究的细菌种类发表的论文数量降低了60%(图1D)。当我们的很多理解来自少量的小动物时,我们对细菌多样性的看法就会有偏见。微生物学家杰弗里·格拉尼克(Jeffery Gralnick)曾经打趣说:“大肠杆菌是大肠杆菌的伟大模型生物。”格拉尼克(Gralnick)的评论提到在Shewanella Oneidensis的TCA周期中发现异常(相对于大肠杆菌)(Brutinel and Gralnick 2012)。尽管Oneidensis链球菌的引用减少了201倍,但可以说不是一个研究的物种。我们的分析将其排名为研究最多的细菌,在所有物种中排名前2.17%。即使是格拉尼克上述论文的简介也将S. oneidensis表示为“模型环境有机体”。如果在微生物2%之外发现了S. Oneidensis的TCA周期等差异,请想象其他98%的微生物中的多样性。微生物学家如何赶上爆炸的生命树?我们提出了两个宏伟的挑战,以培训一代可以解决微生物世界多样性的微生物学家。首先,我们需要采用多因素实验设计。一次进行一次研究的物种,菌株,基因,环境,压力源和表型。统计学家已经教导了数十年来,最有效,最强大的实验设计同时改变了多个因素,然后对效果进行解析

数字 SAT 样题及解释
干扰项解释:选项 A 不正确,因为尽管文章表明 O'Brien 和 Ahmed 得出的结论与其他研究人员不同,但它并没有使用 O'Brien 和 Ahmed 的研究来批评这些早期研究的开展方式或直接质疑这些研究结果的准确性。选项 B 不正确,因为尽管文章表明研究结果发现姿势和认知之间存在联系被夸大了,但它没有提供任何证据表明 O'Brien 和 Ahmed 的研究经常被误解。选项 C 不正确,因为文章表明尽管 O'Brien 和 Ahmed 对研究姿势和认知问题很感兴趣,但它并没有表明这些研究人员在进行研究之前的想法,也没有表明研究人员专门着手解决某个问题。

髓样乳腺癌情况说明书
髓样癌的治疗方法与其他类型的乳腺癌相似。与所有类型的乳腺癌一样,髓样乳腺癌的某些特征会影响可能提供的治疗方法。乳房手术通常是乳腺癌的首选治疗方法。这可能是保乳手术(通常称为广泛局部切除术或乳房肿瘤切除术),即切除癌症及其周围正常乳腺组织的边缘(边界),或乳房切除术(切除包括乳头区域在内的所有乳腺组织)。切除的组织量取决于癌症的大小和乳房的大小。您的乳房外科医生将与您讨论这个问题。