XiaoMi-AI文件搜索系统
World File Search System被子植物
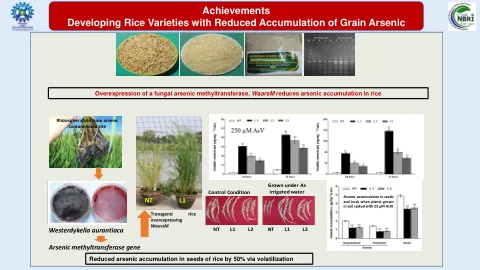
成就开发水稻品种,谷物砷的积累降低
•发现了48种新的地衣和植物,并报告了印度的第一个物种。•调查了23个州和50个受保护区,包括Chambal,Corbett,Gowind WLS,Khaziranga,Kishanpur,Suhelwa,Pachmarhi。•修订了26个分类学复杂或有趣的分类单元。•出版了9个地衣清单和不同地区的植物清单。•书“北方邦的植物资源 - 清单”,其中包括所有藻类,地衣,苔藓植物,孢子菌素,体育植物和被子植物的完整列表。•已推出了北方邦的电子植物。该研究所的植物标本室LWG被国家生物多样性管理局(NBA)公认为“国家存储库”。•启动了植物标本室的数字化,并推出了虚拟标本室。在过去的5年中,植物标本室有15,450个标本,总共3359人参观了植物标本室。

草药DNA降解 - 掉落到非 - ...
使用代表10个被子植物家族的56个基因组敏捷的植物标本室DNA术语,发现重叠的读取对发生在大约80%的读取对中。合并了这种重叠对后,所得的片段及其长度分布被认为反映了实际的DNA碎片。类似于古代DNA中的发生,我们发现在标本室材料中碎片末端的嘌呤过分占代表性。碎片长度的分布适合伽马而不是指数分布,而与标本年龄显然相关。观察到的伽马分布将表明高阶降低动力学,这意味着在降解过程中起作用多种过程。可能,与非重复的植物基因组相比,此处使用的基因组掠夺数据,其中重复序列或隔室具有过多的代表性,具有偏见的基因组片段长度分布和半衰期,但没有可用的数据可用于检验该假设。总体而言,我们的结果表明,我们无法确认是否存在植物档案DNA半寿命以及其速率是多少。

C1a半胱氨酸 - 蛋白酶及其在植物中的抑制剂。
植物半胱氨酸 - 蛋白酶(Cysprot)代表一种良好的蛋白水解酶类型,该酶履行严格调节的生理功能(衰老和种子发芽等)和防御作用。本文集中于帕帕因 - 蛋白酶蛋白酶C1a(Family C1,CA氏族)及其抑制剂植物囊蛋白(Phycys)。尤其是,审查了蛋白酶抑制剂的相互作用及其在整个植物一生中的特定途径的相互参与。c1a cysprot和phycys已被分子表征,比较序列分析已鉴定出共有的功能基序。可以在被子植物中已识别的Cysprot和Phycys数量之间建立相关性。因此,进化力可能已经确定了囊蛋白在这些物种中内源性和害虫性蛋白酶上的控制作用。用荧光蛋白标记蛋白酶和抑制剂揭示了在瞬时转化的洋葱表皮细胞中内质网网络中亚细胞定位的常见模式。通过双分子荧光互补证明了进一步的体内相互作用,这表明它们参与了相同的生理过程。

非防御素肽NPA1在拟南芥中吸引花粉管
摘要在被子植物中,女配子植物分泌了一系列吸引剂,以吸引花粉管进行施肥。在双子蛋白酶中,所有确定的吸引剂都是防御素样半胱氨酸的肽(CRPS)家族成员,而Gramineae中的Zea Mays(如Gramineae中的Zea Mays)使用非CRP型鸡蛋膜1类样肽作为花粉管吸引者。但是,dicots是否具有非Crp吸引剂尚不清楚。在这里,我们表征了拟南芥中非防御素肽诱人的非防御素肽1(NPA1)。NPA1在协同中受MyB98的转录调节。除了特定的花粉管外,NPA1还能够吸引姊妹物种的花粉管A. Lyrata和C. Rubella,但不能吸引E. salsugineum。此外,当引入NPA1以补充MYB98时,它会将花粉管的吸引力和生育能力恢复到与诱饵互补相媲美的水平。一起,这项研究确定了在dicot中的一种新型的肽吸引剂,并突出了吸引提示和信号通路的多样性。

花卉性状的定量遗传分析显示了当前的局限性,但野生的潜在演变
跨被子植物的花卉特征的巨大变化通常被解释为适应授粉媒介的结果。但是,在野生人群中的研究通常没有发现授粉媒介介导的花的迹象。进化理论预测,这可能是稳定条件下停滞期的结果,其次是授粉媒介变化时期较短,为创新表型提供了选择。我们询问停滞周期是否是由于选择稳定,其他形式的选择或低性状能力,即使存在选择,也引起了停滞。我们研究了一种植物,主要是由其范围内的一种蜜蜂授粉的植物。,我们使用了大型野生人群中的全基因组相关性测量性状的遗传力和发展性,并将其与对同一个体的选择估计相结合。我们发现了稳定选择和低性状遗传力的证据,作为流动中停滞的潜在解释。标准花瓣的面积正在稳定下,但可变性不是可遗传的。单独的特征,花的重量具有很高的含义,但目前尚未选择。我们展示了一个简单的授粉环境如何与当前的自适应进化变化前提条件相吻合,而遗传性的变化仍在响应未来的选择压力。

Asteraceae基因组学的综合平台
Asteraceae是最大的被子植物家族,因其出色的药用,园艺和观赏价值引起了广泛的关注。然而,关于星形科植物的研究由于复杂的遗传背景而面临挑战。随着测序技术的持续发展,从星状科物种中积累了大量的基因组和遗传资源。这促使对这个多样化的植物群中对全面的基因组分析的需求。为了满足这种需求,我们开发了Asteraceae基因组学数据库(AGD; http://cbcb.cdutcm.edu.cn/agd/)。AGD充当集中和系统的资源,赋予了各种领域的研究人员,例如基因注释,基因家族分析,进化生物学和遗传育种。AGD不仅包含高质量的基因组序列和细胞器基因组数据,而且还提供了广泛的分析工具,包括BLAST,JBROWSE,SSR FINDER,HMMSEARZER,HMMSEARCH,HEMMAP,HEATMAP,PRIMER3,PLANTIMSISMASH和CRISPRCASFINDER。这些工具使用户能够方便地查询,分析和比较各种星际科中的基因组信息。AGD的建立在推进Asteraceae基因组学,促进遗传育种并通过为研究人员提供全面且用户友好的基因组资源平台来维护生物多样性方面具有巨大的意义。

黑麦的遗传冗余以多种方式显示
摘要:五十年前,苏苏姆·ohno(Susumu Ohno)提出了著名的C值悖论,该悖论指出,基因组的物理大小,即DNA的量与生物体的复杂性之间没有相关性,并突出了基因组降低的问题。DNA已被描述为“垃圾或selfer dNA”。垃圾DNA的有争议的概念仍然可行。rye是对该概念的正确性和科学意义的另一个测试的便捷主题。栽培黑麦的基因组,塞莱·瓦雷·L。被认为是部落小毛虫的物种中最大的一部分之一,因此它是平均被子植物的基因组及其最接近进化邻居的基因组,例如大麦,荷尔德人,荷尔德人(大约30-35%)和二型麦田(Triticum),triticum,triticum,triticum,triticum,triticum of triticum of triticum,triticum,and triticum of diplitium of triticum,and。审查提供了对黑麦染色体各个区域的结构组织的分析,并描述了有助于其在进化过程中大小增加的分子机制以及这些过程中涉及的DNA序列的类别。是真核基因组冗余概念发展的历史,并讨论了此问题的当前状态。

茎分生组织的细胞间通讯
大多数被子植物的茎尖分生组织 (SAM) 呈圆锥形,由高度组织化的细胞层和功能域组成(111)(图 1)。最外层(L1)产生表皮组织,下一层(L2)产生表皮下组织和配子。L1 和 L2 都通过垂周细胞分裂保持为离散的细胞层,由此形成垂直于分生组织表面的新细胞壁,而子细胞则留在其原始层中。因此,从遗传学上讲,L1 和 L2 是克隆。体细胞突变由子细胞遗传,子细胞将保留在同一细胞层中,从而产生嵌合植物组织。分生组织较深区域的细胞形成第三层(L3)。在这里,细胞分裂的方向性较差,L3 产生大部分植物茎组织、维管系统和植物叶片的内层。包括花分生组织在内的新器官原基的生成发生在外周区 (PZ) 中分生组织的侧面,而分生组织的中心由中心区 (CZ) 中未分化且很少分裂的干细胞组成。SAM 和花分生组织 (FM) 具有相同的一般结构,但有一个重要区别:FM 中的干细胞用于

花卉性状的定量遗传分析显示了当前的局限性,但野生的潜在演变
跨被子植物的花卉特征的巨大变化通常被解释为适应授粉媒介的结果。但是,在野生人群中的研究通常没有发现授粉媒介介导的花的迹象。进化理论预测,这可能是稳定条件下停滞期的结果,其次是授粉媒介变化时期较短,为创新表型提供了选择。我们询问停滞周期是否是由于选择稳定,其他形式的选择或低性状能力,即使存在选择,也引起了停滞。我们研究了一种植物,主要是由其范围内的一种蜜蜂授粉的植物。,我们使用了大型野生人群中的全基因组相关性测量性状的遗传力和发展性,并将其与对同一个体的选择估计相结合。我们发现了稳定选择和低性状遗传力的证据,作为流动中停滞的潜在解释。标准花瓣的面积正在稳定下,但可变性不是可遗传的。单独的特征,花的重量具有很高的含义,但目前尚未选择。我们展示了一个简单的授粉环境如何与当前的自适应进化变化前提条件相吻合,而遗传性的变化仍在响应未来的选择压力。

小家庭,重大影响:RNL助手NLR及其在植物先天免疫中的重要性
通量,活性氧的产生和有丝分裂原激活的蛋白激酶激活[1]。最近的研究表明,2受体系统的相互依赖性和相互增强[2,3]。基于其N末端结构域及其系统发育,NLR在盘绕型圈(CC)结构域,Toll-like/interleukin-1受体耐药性(TIR)结构域中被构成,对白粉病(CC R)的耐药性(CC R)域的耐药性包含NLR,含有NLR,含有AS CNLS,TONLS,the and cnls for and thls for and for and thls from thls&tnls for and。在拟南芥中(以下称为Arabidopsis),多个PRR和效应子传感NLR(某些CNL和所有测试的TNL)需要存在RNL,也称为Helper NLR,以激活全部免疫力[5,6]。rnls形成一个由2个亚家族组成的小而进化保守的进化枝,活化的抗耐药性1(ADR1)和N需求基因1(NRG1)家族,它们在血管植物的发散之前已有分离[4]。拟南芥基因组径流3 ADR1和2 NRG1全长基因需要完全免疫[7-9]。尽管RNL仅代表大多数被子植物中NLR基因库的一小部分[4,10],但对于植物而言,它们至关重要。在这里,我们重点介绍了RNL在免疫过程中的功能以及讨论RNL激活机制的最新发现。