XiaoMi-AI文件搜索系统
World File Search System辅之以

本文综述了结构磁共振成像在常见神经精神类疾病中的应用
[3] LIBOWITZ MR,WEI K,TRAN T,et al.Regional brain volumes relate to Alzheimer's disease cerebrospinal fluid biomarkers and neuropsychometry:A cross-sectional,observational study[J].PLoS One,2021,16(7):e0254332.[4] 王含春 , 汪群芳 , 罗长国 , 等 .磁共振薄层扫描结合人工智能脑结构分割技术分析海马体积辅 助诊断脑小血管病认知功能障碍 [J].全科医学临床与教育 ,2024,22(3):208-211.[5] 姜华 , 宛丰 , 吕衍文 , 等 .2 型糖尿病伴认知功能障碍患者基于体素的脑形态学 MRI 研究 [J].中 国 CT 和 MRI 杂志 ,2018,16(4):22-25.[6] 景赟杭 , 郭瑞 , 常轲 , 等 .2 型糖尿病性认知功能障碍脑结构 MRI 成像研究进展 [J].延安大学学 报(医学科学版) ,2024,22(1):88-91,107.[7] 郭浩 , 和荣丽 .磁共振成像对老年性痴呆患者海马解剖结构的评估价值研究 [J].磁共振成 像 ,2022,13(8):75-79.[8] 罗财妹 , 李梦春 , 秦若梦 , 等 .阿尔茨海默病谱系患者的海马亚区体积损害特征 [J].中风与神经 疾病杂志 ,2019,36(12):1097-1101.[9] 冯伦伦 , 金蓉 , 曹城浩 , 等 .阿尔茨海默病患者认知功能减退的海马亚区结构改变分析 [J].临床 放射学杂志 ,2022,41(10):1819-1824.[10] WEI Y,HUANG N,LIU Y,et al.Hippocampal and Amygdalar Morpho logical Abnormalities in Alzheimer,s Disease Based on Three Chinese MRI Datasets[J].Curr Alzheimer Res,2020,17:1221-1231 . [11] ESTEVEZ S S,JIMENEZ H A,ADNI G.Comparative analy sis of methods of volume adjustment in hippocampal volumetry for the diagnosis of Alzheimer disease[J].Neuroradiol,2020;47(2):161-5.[12] 曾利川 , 王林 , 廖华强 , 等 .结构与功能磁共振成像在轻度认知障碍及阿尔茨海默病中的应 用 [J].中国老年学杂志 ,2021,41(13):2902-2907.[13] KODAM P,SAI S R,PRADHAN S S,et al.Integrated multi-omics analysis of Alzheimer's disease shows molecular signatures associated with disease progression and potential therapeutic targets[J].Sci Rep,2023,13(1):3695.[14] 黄建 , 王志 .复杂网络分析技术在阿尔兹海默症患者脑结构和功能影像中的应用进展 [J].中 国医学物理学杂志 ,2024,41(8):1053-1055.[15] JELLINGER K A.The pathobiological basis of depression in Parkinson disease:challenges and outlooks[J].J Neural Transm(Vienna),2022,129(12):1397-1418.[16] BANWINKLER M,THEIS H,PRANGE S,et al.Imaging the limbic system in Parkinson's disease-A review of limbic pathology and clinical symptoms[J].Brain Sci,2022,12(9):1248.[17] 程秀 , 张鹏飞 , 王俊 , 等 .小脑结构与功能磁共振成像在帕金森病中的研究进展 [J].磁共振成 像 ,2022,13(4):146-149.[18] CUI X,LI L,YU L,et al.Gray Matter Atrophy in Parkinson's Disease and the Parkinsonian Variant of Multiple System Atrophy:A Combined ROI-and Voxel-Based Morphometric Study[J].Clinics(Sao Paulo),2020,75:e1505.[19] LOPEZ A M,TRUJILLO P,HERNANDEZ A B,et al.Structural Correlates of the Sensorimotor Cerebellum in Parkinson's Disease and Essential Tremor[J].Mov Disord,2020,35(7):1181-1188.[20] 鲍奕清 , 王二磊 , 邹楠 , 等 .帕金森病伴疲劳患者的大脑功能与结构磁共振成像研究 [J].临床 放射学杂志 ,2024,43(8):1265-1270.[21] 邹楠 , 王二磊 , 张金茹 , 等 .帕金森病伴疼痛患者大脑皮层厚度改变的结构 MRI 研究 [J].磁共 振成像 ,2024,15(5):13-18,23.[22] 屈明睿 , 高冰冰 , 苗延巍 .帕金森病伴抑郁在脑边缘系统结构及功能改变的 MRI 研究进展 [J].磁共振成像 ,2023,14(12): 127-131.

同种型柠檬酸可治愈肺炎克雷伯菌的 NifV 表型
固氮酶催化 N2 还原为铵 (1)。固氮酶由两种蛋白质组成,即二氮酶 (组分 I,Mo-Fe 蛋白) 和二氮酶还原酶 (组分 II,Fe 蛋白) (1, 3)。二氮酶含有一个独特的辅基,即铁钼辅因子 (FeMo-co),由 Fe、Mo 和 S (15) 组成。生化和遗传研究表明,至少有六种 nif (固氮) 基因产物参与了 FeMo-co 的生物合成。含有 nifB、nifN 或 nifE 突变的肺炎克雷伯菌菌株无法合成 FeMo-co (12, 15)。在含有低水平钼酸盐的培养基中,当固氮酶被解除抑制时,nifQ 突变的菌株不会合成 FeMo-co (8)。某些含有 nifH(编码二氮酶还原酶)突变的肺炎克雷伯菌和棕色固氮菌菌株无法积累 FeMo-co(2, 13)。从含有 nifV 突变的肺炎克雷伯菌菌株中分离出的二氮酶表现出改变的底物亲和力和抑制剂敏感性(10)。进一步的研究表明,NifV 突变体在 FeMo-co 合成方面存在缺陷(4)。最近,描述了一种体外合成 FeMo-co 的系统,该系统需要 ATP、钼酸盐、nifB、nifN 和 nifE 的基因产物(17)、二氮酶还原酶(未发表的数据)和同型柠檬酸(6)。肺炎克雷伯菌对同型柠檬酸的积累与功能性 nifV 基因的存在有关,该基因显然编码同型柠檬酸合酶(7)。在解除固氮酶抑制期间,发现高柠檬酸在肺炎克雷伯氏菌培养物培养基中积累 (6)。我们在此报告,向肺炎克雷伯氏菌 NifV 突变体培养基中添加高柠檬酸可治愈该表型。肺炎克雷伯氏菌 UN 是从菌株 M5al 中重新分离的野生型菌株,该菌株最初来自 PW Wilson 的收藏。菌株 UN1991 (nifV4945) 是一种稳定的 NifV 突变体,回复频率为 3 x 10-10(T. MacNeil,博士论文,威斯康星大学麦迪逊分校,1978 年),之前已有描述 (9)。肺炎克雷伯氏菌突变体中的生长和固氮酶解除抑制已被描述 (8)。从肺炎克雷伯菌 (6) 培养物的去阻遏培养基中分离出 (R)-2-羟基-1,2,4-丁烷三羧酸 (高柠檬酸)。将高柠檬酸添加到 UN1991 培养物中,最终浓度约为 83 mg * 升-' (0.4 mM)。用 DEAE-纤维素色谱法 (14) 从菌株 UN、UN1991 和 UN1991 中纯化二氮酶,这些菌株在高柠檬酸存在下已对固氮酶进行了去阻遏。已描述了乙炔和 N2 还原测定

CP研究新闻2024- 9月2日
1。Association between upper limb clinical tests and accelerometry metrics for arm use in daily life in children with unilateral cerebral palsy Jenny Hedberg-Graff, Lucian Bezuidenhout, Lena Krumlinde-Sundholm, Jenny Hallgren, David Moulaee Conradsson, Maria Hagströmer Disabil Rehabil.2024年8月27日:1-7。 doi:10.1080/09638288.2024.2393801。在线印刷前线。目的:评估单侧脑瘫(CP)儿童的上限(UL)临床测试与UL加速度测定指标之间的关联。方法:在这项横断面研究中,包括二十名单侧CP和手动能力分类系统I-III的儿童。在临床环境和日常生活中收集了辅助手评估,框和加速度测定指标的结果。UL不对称指数(即,功能良好的UL与受影响的UL使用之间的比率),并在日常生活中评估了UL的相对使用。Spearman的相关性用于确定临床环境和日常生活中UL临床测试与加速度测定指标之间的关联。结果:在日常生活中久坐时间(= -0.64)期间,辅助手部评估单元与加速度计指标之间的最强负相关性。与儿童在光线下(不对称指数:23.97)或更高的强度体育活动(不对称指数:不对称指数:13.39)相比,ULS之间的不对称性在儿童久坐时间(不对称指数:45.15)中最高(不对称指数:45.15)。在日常生活中,孩子们同时使用了44%的时间。结论:通过量化UL运动量和上肢之间的不对称量,加速度计指标可以为临床测试提供其他客观信息。pmid:39192545 2。Effect of touch screen tablet use on fine motor functions in children with hemiparetic cerebral palsy: A randomized controlled trial Hanaa M Abd-Elfattah, Dina O Shokri M Galal, Shaima M Abdelmageed, Sobhy M Aly, Fairouz H Ameen, Asmaa O Sayed, Amira M Abd-Elmonem Randomized Controlled Trial NeuroRehabilitation.2024; 55(1):137-146。 doi:10.3233/nre-240134。背景:脑瘫是影响中枢神经系统并引起大量残疾的最常见状况。目的:确定触摸屏片剂对偏瘫儿童的精细运动功能的影响。方法:这是一项随机对照试验,涉及60名儿童,年龄从5至7岁不等,随机分为两组:干预或对照组(每组30个儿童)。两组连续12周进行了设计的精细运动任务。此外,在三十分钟内,干预组在触摸屏平板电脑上获得了精美的运动锻炼程序。上肢功能,手指敏捷性和捏合强度分别使用上肢技能测试(QUEST),九孔PEG测试和Jamar液压捏量规来测量建议的治疗计划。结果:入院时干预组之间所有结果度量均等效(p> 0.05)。在两组内的所有评估变量中都发现了显着改善。同时,干预组有

2024 年会议
海报展示 1 49 (PO-01) Igor Varga - 自动颅骨缝合线检测用于小鼠表型分析 51 (PO-02) Michaela Šímová - 揭示小鼠卵黄囊中红细胞和髓系祖细胞的出现 52 (PO-03) Olha Pyko - 揭示 ZNF644 缺失的影响:研究 C2H2 锌指蛋白在小鼠雌性表型中的作用 53 (PO-04) Rodolfo Favero - 开发和鉴定 Netherton 综合征的条件性 Spink5 基因敲除小鼠模型 54 (PO-05) Hirotoshi Shibuya - 使用新型增强微型 CT 开发高通量、高分辨率软组织成像方法 55 (PO-06) Matilde Vale - 开发用于治疗钻石的治疗性外泌体和基因疗法黑粉病 (DBA) 56 (PO-07) Sabina Cerulová - 最初创建的具有罕见 GALNT3 突变的小鼠模型中钙磷酸代谢失调 57 (PO-08) Zhenni Liu - 探索 GPR45 在代谢调节中的作用及其对肥胖和相关疾病的影响 58 (PO-09) Eni Tomovic - 在捷克儿科患者中检测到的 GRIN 变异的遗传和功能分析 59 (PO-10) Ben Davies - Grem1 (88 kb) 和 Taf1 (166kb) 基因的人类基因组人源化 60 (PO-11) Federica Gambini - 用于 SARS-CoV-2 研究的新型可诱导 hACE2 小鼠模型的表征:对急性感染和长期 COVID 的见解 61 (PO-12) Klevinda Fili - 携带神经发育疾病相关变异的小鼠的表征62 (PO-13) Vera Abramova - 敲除 NMDA 受体 grin2Aa 和 grin2Ab 基因的斑马鱼幼虫的特征:基因表达和游泳行为 63 (PO-14) Hana Kolesová - Jagged1 条件性缺失和基于患者的单一变体小鼠模型的形态学和生理学 64 (PO-15) Petr Nickl - AAV 载体在小鼠植入前胚胎中进行多步等位基因转换 65 (PO-16) Silvia Mandillo - 肌肉特异性基因编辑改善了 1 型肌强直性营养不良小鼠模型中的分子和表型缺陷 66 (PO-17) Kristýna Neffeová - 法洛四联症小鼠模型中 Jagged1 缺失的生理和形态学后果 67 (PO-18) Tomasz Kowalczyk - 蛋白质组学PACS2 基因突变小鼠软组织的分析 68 (PO-19) Dominik Cysewski - PACS2 E209K 突变小鼠脑组织的蛋白质组学和代谢组学分析:深入了解分子失调 69 (PO-20) Betul Melike Ogan - FAM83H 在免疫系统稳态中的作用 70 (PO-21) Maximilián Goleňa - C57Bl/6NCrl 小鼠测量参数的季节性 71 (PO-22) Tobiáš Ber,Kateryna Nemesh - 陆生蛞蝓作为研究 RNA 沉默途径的潜在动物模型 72 (PO-23) Gunay Akbarova-Ben-Tzvi - 修饰的 TGF-β β 家族对整合素-ββ1 合成软骨细胞片的影响 73 (PO-24) Arkadiusz Żbikowski - PACS2 综合征对小鼠肺和肾结构的影响 75 (PO-25) Viktor Kostohryz - 附加基因治疗的前景 76 (PO-26) Miles Joseph Raishbrook - Fam84b 在视网膜稳态中的重要性 77 (PO-27) JI XU - 转录辅阻遏物 TLE1 是脂肪细胞分化的积极因素 78 (PO-28) Sylvie Dlugosova - 骨骼畸形和矿化缺陷Fgf20 KO 小鼠 79
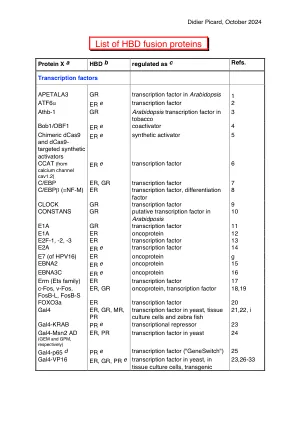
HBD 融合蛋白列表 - Picard 实验室 |
Picard, D. (2000)。通过与类固醇结合域融合实现蛋白质的翻译后调控。Methods Enzymol. 327 , 385-401。ER α HBD 融合在小鼠中的应用:Whitfield, J.、Littlewood, T.、Evan, GI 和 Soucek, L. (2015)。小鼠模型中的雌激素受体融合系统:可逆转换。Cold Spring Harb. Protoc. 2015 , 227-234。参考文献 1. Sablowski, RW 和 Meyerowitz, EM NO APICAL MERISTEM 的同源物是花同源基因 APETALA3/PISTILLATA 的直接靶标。Cell 92 , 93- 103 (1998)。 2. Thuerauf, DJ, Marcinko, M., Belmont, PJ 和 Glembotski, CC ATF6α 和 ATF6β 异构体特异性特征对内质网应激反应基因表达和细胞活力的影响。J. Biol. Chem. 282, 22865-22878 (2007)。3. Aoyama, T. 等人。拟南芥转录激活因子 Athb- 1 的异位表达改变了烟草叶细胞的命运。Plant Cell 7, 1773-1785 (1995)。4. Laumen, H., Nielsen, PJ 和 Wirth, T. BOB.1/OBF.1 辅激活因子对 B 细胞中八聚体依赖性转录至关重要。Eur. J. Immunol. 30, 458-469 (2000)。 5. Lu, J. 等人。用于转录激活和基因组编辑的多模式药物诱导 CRISPR/Cas9 装置。核酸研究。46,e25(2018)。6. Gomez-Ospina, N.、Tsuruta, F.、Barreto-Chang, O.、Hu, L. 和 Dolmetsch, R. L 型电压门控钙通道 ca(v)1.2 的 C 端编码转录因子。细胞 127,591-606(2006)。7. Umek, RM、Friedman, AD 和 McKnight, SL CCAAT 增强子结合蛋白:分化开关的组成部分。科学 251,288-292(1991)。 8. Müller, C., Kowenz-Leutz, E., Grieser-Ade, S., Graf, T. & Leutz, A. NF-M(鸡 C/EBP beta)诱导造血祖细胞系嗜酸性分化和凋亡。EMBO J. 14 , 6127-6135 (1995)。9. McDonald, MJ & Rosbash, M. 果蝇昼夜节律基因表达的微阵列分析和组织。Cell 107 , 567-578 (2001)。10. Simon, R., Igeno, MI & Coupland, G. 拟南芥花分生组织身份基因的激活。Nature 384 , 59-62 (1996)。 11. Picard, D., Salser, SJ & Yamamoto, KR 糖皮质激素受体类固醇结合域内可移动且可调节的失活功能。Cell 54 , 1073-1080 (1988)。12. Spitkovsky, D. 等人。腺病毒 E1A 在具有 E1A 依赖性条件性增殖的细胞系中对细胞周期蛋白基因表达的调节。J. Virol. 68 , 2206-2214 (1994)。13. Vigo, E. 等人。CDC25A 磷酸酶是 E2F 的靶标,是 E2F 有效诱导的 S 期所必需的。Mol. Cell. Biol. 19 , 6379-6395 (1999)。 14.Jones, ME, Kondo, M. & Zhuang, Y. A tamoxifen inducible knock-in allele for investigation of E2A function. BMC Dev. Biol. 9 , 51 (2009). 15.Zhao, B. et al. RNAs induced by Epstein-Barr virus Nuclear Antigen 2 in Lymphoblastoid cell lines. Proc. Natl. Acad. Sci. USA 103 , 1900-1905 (2006). 16. Maruo,S. 等人。Epstein–Barr 病毒核蛋白 EBNA3C 是淋巴母细胞细胞周期进程和生长维持所必需的。美国国家科学院院刊 103,19500-19505(2006 年)。
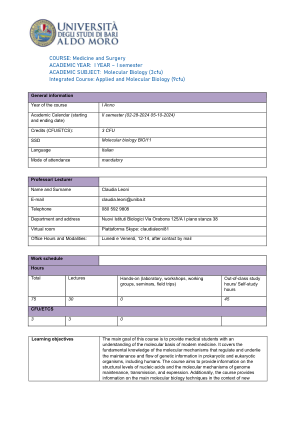
分子生物学(3cfu)综合课程
超螺旋和拓扑性质。拓扑异构酶。细菌类核。组蛋白和核小体的性质和组装。染色质的高级结构。组蛋白的翻译后修饰。溴多胺和染色质结构域。表观遗传学。原核生物和真核生物的基因组。复制模型。DNA合成。细菌DNA聚合酶。校对和缺口翻译。复制子模型。OriC和半甲基化。Ter/Tus。真核细胞核中的复制工厂。ARS结构和复制控制。酶学。前RC和前启动复合物。复制抑制剂,如化疗药物和抗病毒药物。端粒和端粒酶的结构、功能和意义。DNA损伤和修复。基因组作为动态实体。体细胞和种系突变。SNP。内在和外在损伤。化学和物理诱变剂。原核生物和真核生物中的去除、逆转和损伤避免系统。MUT 系统。BER 系统。糖基化酶的重要性。安全系统。NER 系统:UvrABCD 和 XP 蛋白。GG-NER 和 TC-NER。光解作用、MGMT、AlkBH。损伤耐受机制。TLS。细菌中的 SOS 反应。单丝和双丝断裂。HR 和 NHEJ。由于修复系统突变而导致的人类疾病。位点特异性重组。重组酶。Lambda 噬菌体。Cre-Lox 系统和 KO 小鼠。简单和复杂的转座子。SINE 和 LINE 元素、Alu 序列。原核生物和真核生物中的 RNA。结构、类型和特性。细菌 RNA 聚合酶和相关因子。转录单位。转录步骤。细菌启动子中的共识序列。终止机制。抑制剂。 Lac、ara 和 trp 操纵子。阳性和阴性对照。真核细胞中的 RNA 类别。RNA 聚合酶 (CTD) 的结构和功能。三种启动子的特征。基础转录机制。TFIIH。反式激活因子、辅激活因子。CpG 岛甲基化。组蛋白密码。长程调节剂。DNA 结合蛋白的功能域 (HTH、HD、HLH、ZF、LZ)。RNA 成熟、核运输和转录后控制。加帽类型。添加 polyA。CTD 的变化。外显子和内含子。外显子改组。四类内含子及其去除机制。剪接体和剪接位点。AT-AC 剪接。EJC 复合体。可变剪接。ESE 和 ESS 序列、SR 和 hnRNP 蛋白。SMN 基因。剪接和病理。rRNA 和 tRNA 加工反应。核糖体基因。 SnoRNA 和核仁功能。RNA 编辑。插入和转换编辑。人类 RNA 编辑的示例。细胞核和细胞质中的 RNA 周转。外泌体。无义介导的 mRNA 衰变 (NMD)。非编码 RNA。小 RNA 在细胞中的功能。RNA 干扰。siRNA。微小 RNA 的生物发生。miRNA、长链非编码 RNA、环状 RNA 的作用机制。逆转录病毒的一般信息。遗传密码和翻译。遗传密码的性质和特征。线粒体密码。ORF。tRNA 的特征。不常见碱基。aa-tRNA 合成酶的功能和类别。遗传密码的翻译重编码和扩展。SeCys。核糖体是一种核酶。原核生物和真核生物的翻译阶段。不同的启动机制。能量成本。NSMD。细菌中的 tmRNA。抑制剂。蛋白质的翻译后修饰、分选和降解。折叠和错误折叠。朊病毒。HSP60 和 HSP70。泛素和泛素化系统。SUMO 化糖基化。蛋白酶体。肽信号。蛋白质分选。线粒体输入。线粒体基因组细胞中的线粒体可塑性。人类线粒体基因组。遗传、结构、复制及其表达的原理。线粒体 DNA 中的改变。DNA 克隆的原理。修饰限制系统。克隆载体。cDNA 合成。基因组 DNA 和 cDNA 文库。TA 克隆。表达克隆。基因表达沉默。基因治疗。数据库。基因组编辑元件(Talen、Zn 指、CRISPR/Cas9 系统)。PCR 和 DNA 测序。PCR 的特性。PCR-RFLP。实时 PCR、DNA 测序。NGS。核酸杂交。杂交原理。熔点和严格性。探针制备:切口平移。Southern、Northern、杂交测定。蛋白质印迹。

散热器
基因工程是指对基因结构的操纵或改变,以在生物体中产生所需的特征。此过程涉及破坏和连接DNA分子,以及从一种物种将基因移植或剪接基因进入另一种宿主物种。如果添加来自其他物种的遗传物质,则可以称为转基因。基因工程主要集中于操纵遗传物质(DNA)以预定的方式实现特定目标。这可能涉及更改一个碱基对(A-T或C-G),删除DNA的整个区域,或引入基因的其他副本。它也可能涉及从另一生物的基因组中提取DNA,并将其与个人自己的DNA结合。通过基因工程改变的植物,动物或微生物被称为转基因生物(GMO)。如果将来自另一种物种的遗传物质添加到宿主中,则该术语适用于宿主。Cisgenic是指使用可以自然与宿主繁殖的物种中的遗传物质,而当从靶向生物中去除遗传物质时,敲除生物会产生敲除生物。基因工程的历史可以追溯到1970年代,杰克·威廉姆森(Jack Williamson)在他的科幻小说小说《龙岛》(Dragon's Island)中首先创造了一词。赫伯特·博耶(Herbert Boyer)和斯坦利·科恩(Stanley Cohen)在1973年将抗生素抗性基因插入大肠杆菌细菌中创建了第一个转基因生物。1986年在法国和美国对第一批基因工程植物进行了测试,烟草植物设计为具有抗除草剂的抗性。1。2。基因工程的应用包括科学研究,农业和技术。在植物中,它提高了土豆,西红柿和大米等农作物的韧性,营养价值和生长速度。在动物中,它已被用来开发在其牛奶中产生治疗蛋白的绵羊,以治疗囊性纤维化,或者在黑暗中发光的蠕虫。遗传工程可用于从目标生物体中去除遗传物质,从而产生敲除生物。此过程涉及操纵DNA分子以实现特定目标,并在各个领域具有深远的影响。允许科学家通过了解遗传因素来研究像阿尔茨海默氏症这样的疾病。转基因的生物用于农业,医学和其他领域。其中包括已设计为具有理想性状或特征的转基因植物,动物,甚至人类。此类生物的例子包括Flavr Savr番茄,BT-COTTON,金米,蓝色玫瑰,发光鱼和绵羊Dolly。基因工程涉及使用各种工具和技术修改生物体的DNA。这些工具(称为分子剪刀和分子胶)用于切割和连接DNA序列,使科学家可以引入新基因或修改现有基因。在产生胰岛素,酵母和细菌的情况下。大肠杆菌经过基因设计以产生类似人类的胰岛素,后来批准用于糖尿病患者。然后将所得的胰岛素纯化并包装以分配。3。4。5。6。此过程涉及多个步骤,包括从细菌中提取质粒DNA,使用限制酶切割质粒,将其插入人类胰岛素的基因,将修饰的质粒引入新细胞中,并生长这些细胞以产生大量的胰岛素。遗传工程师还利用分子工具,例如限制性核酸内切酶,在特定位置切割DNA和DNA连接酶,将DNA片段融合在一起。**分子剪刀:限制位点**限制位点,也称为分子剪刀,是具有特定点的DNA分子,可以使用限制性酶切割双链DNA。**生物学作用和防御机制**大多数细菌都使用限制酶来防御噬菌体(感染细菌的病毒)。这些酶通过将其DNA在特定部位切割,以甲基DNA在腺嘌呤或胞嘧啶碱基中保护宿主DNA来防止噬菌体复制。**限制酶的历史**第一个限制酶在1970年由Hindlil分离出来。从那时起,已经研究了超过3000个酶,并且有600多种可用于DNA修饰和操纵。**作用机理**限制性核酸内切酶扫描DNA的长度,与特定序列结合,并通过水解磷酸二酯键在双螺旋的每个糖磷酸骨架中切成一个切割。**限制片段的类型**限制酶产生两种类型的切割:钝的末端和粘性末端。钝器末端可以连接到任何其他带有钝端的DNA碎片,而粘性末端可以结合起来从不同来源创建新分子。7。**交错的切割和粘性末端**大多数限制性酶会产生交错的切割,产生单链的“粘性末端”。这些粘性末端是互补的,可以从不同来源创建和操纵DNA序列。**限制性酶**限制性核酸内切酶分为三个一般组:I型,II型和III型,基于其组成,酶辅因子的需求,靶序性性质和DNA裂解位点相对于目标序列。在这里给出的文本•基因工程通过允许对遗传物质进行精确修改,从而显着影响了医学,取证和农业领域。•选择性育种涉及在生物体中选择特定特征以传递到其后代。•基因剪接可以使用实验室技术(例如PCR)故意改变DNA序列。•克隆涉及通过重复的PCR过程创建多个基因的副本,然后将其插入其他DNA链中以产生蛋白质。•可以通过将基因从一个生物体移植到另一种生物来创建遗传修饰的生物(GMO),从而导致以前不存在新的特征。•转基因生物的例子包括太阳托里的“蓝色”玫瑰,产生一种用于血液凝血疾病的稀有蛋白质的山羊,以及为不足者提供维生素补充维生素的金米。转基因的生物:GMO,基因疗法,干细胞,克隆和取证DNA指纹的概述揭示了样本之间的相似性,有助于证明或建立家庭关系,而人类基因组项目则解释了人类DNA,以了解人类DNA,以了解疾病和推动各种领域的科学突破,并在各种领域中驾驶,并在各种领域中进行效果。