XiaoMi-AI文件搜索系统
World File Search System酪乳

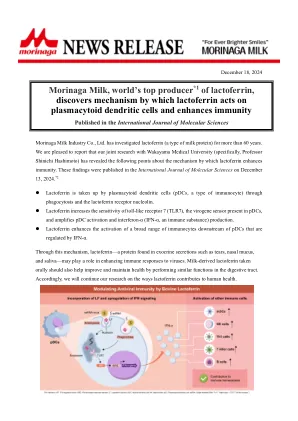
乳酸化修饰在年龄相关性疾病中的研究新进展
随着对乳酸化研究的不断深入,蛋白质乳酸化修饰 越来越受到研究者的关注。而乳酸生成及代谢异常、基 因表达、修饰串扰等因素影响着乳酸化修饰动态平衡过 程。乳酸化修饰不仅在正常的细胞活动中发挥重要作用, 也参与调控年龄相关性疾病的发病机制。组蛋白乳酸化 主要通过调节相关基因的转录和表达来影响细胞的功能 状态,非组蛋白乳酸化则可以通过促进EndoMT,激活 信号通路,亚细胞定位和翻译后修饰串扰等功能,导致 年龄相关性疾病的发生和发展。然而,乳酸化修饰的调 控机制的研究尚且处于起步阶段,仍有许多未知功能和 新的修饰酶有待进一步探索,目前这些研究有助于揭示 乳酸化修饰的分布和调控机制以及在多种年龄相关性疾 病中的作用效果,并以此为依据转化为可应用于临床治 疗的手段是亟待解决的问题 。

第13章
肠道微生物在维持整体健康中起着至关重要的作用,尤其是在营养的消化和吸收中。益生元和益生菌在影响肠道健康的各种组成部分中引起了极大的关注。阿育吠陀(Ayurveda),古老的印度医学系统非常重视饮食与健康之间的关系。在阿育吠陀(Ayurveda Takra)(酪乳)中包括在Pathyakalpana(饮食制剂)下,可以用作药物和饮食方案。takra(酪乳)被认为是益生菌富含食物,其中含有有益的细菌,例如乳酸杆菌,可以有助于健康的肠道菌群并支持肠道健康。根据阿育吠陀(Ayurveda Laghu)(灯光),dipana(开胃),格雷(Grahi)(吸收)特性,使其成为支持消化健康的绝佳选择。ardrak(Zingiber officinale),通常称为生姜的药用特性高度重视,尤其是与消化系统有关。根据阿育吠陀的特性,例如dipana(开胃),Pachana(有助于消化),Amapachana(净化毒素)支持消化和免疫系统。Ardrak的抗菌,抗炎和抗氧化特性可以间接支持乳杆菌的作用,并为更健康的肠道环境做出贡献。合并后,塔克(Takra)和生姜被认为可以增强他人对肠道的有益作用。根据普拉克里提(Dosha)(多沙的占主导地位)和特定的消化需求,消费takra和姜很重要。本文探讨了Takra和Ginger对滋养肠道微生物组和促进消化的共生影响。

人乳寡糖:结构和功能
寡糖(来自希腊语ὀλίγοςOlígos,“少数”和σάκχαρSácchar,“糖”)是糖(糖)聚合物,其中包含少量数量(通常为3-10个或更多)单糖(简单糖)。与大多数其他哺乳动物的牛奶不同,人乳是独特的,因为它含有高浓度的150多种不同且结构上不同的寡糖。实际上,对于5-15 g/L,成熟牛奶中的人牛奶寡糖(HMO)的总浓度通常超过人奶蛋白的总浓度,使HMOS成为仅次于简单的牛奶糖乳糖和脂质的第三大分子,而不是计算水[1]。HMO包含多达5个不同的构建块(单糖):葡萄糖(GLC),半乳糖(GAL),N-乙酰基葡萄糖胺(GLCNAC),Fucose(FUC)和唾液酸(SIA)。根据使用了哪些构建块以及如何将它们链接在一起[1],从而生成不同的HMO。图1a显示了HMO结构组件的蓝图。所有HMO在还原端携带乳糖(GALβ1-4GLC)。乳糖可以通过二糖乳糖-N-生物(GALβ1–3GLCNAC)或n-乙酰氨基胺(GALβ1-4GLCNAC)的添加来拉长。乳糖或细长链可以用唾液酸在α2-3-或α2-6-链接中修饰,在α1-2-,α1-3-或α1-3-或α1-4链接中进行葡萄糖基化,从而大大扩展了HMO结构组合的多样性。对于外部,每种唾液酸单糖都包含一个羧基,并引起对HMO分子的负电荷,从而改变了其结构特性。HMO结构通常决定其功能[2]。尽管HMO组成遵循基本的蓝图和150多个不同的HMO,但迄今已确定了150多个不同的HMO,但重要的是要注意,每个女性都合成并分泌出不同的HMO组成曲线,在不同女性之间有很大的不同(图1b),但在同一妇女的哺乳过程中保持相当恒定[3]。到目前为止,我们的实验室已经分析了从世界各地女性收集的10,000多个牛奶样本中的HMO组成,作为各种协作项目的一部分。图1C列出了主成分(PC)图中的某些数据,再次强调了女性之间的HMO组成图谱有所不同,但也存在明显的HMO剖面簇或HMO lactotypes。


无乳链球菌疫苗:现状和...
无乳链球菌是新生儿、老年人和患有合并症的成年人侵袭性感染的主要原因之一。新生儿感染无乳链球菌的主要危险因素是妊娠期间母亲直肠阴道定植,无乳链球菌可导致宫内感染、早产和/或死产 (1)。2020 年,全球约有 2000 万孕妇被这种微生物定植,近 40 万儿童患有早发性无乳链球菌病 (EOD,出生后 0 至 6 天) 或晚发性无乳链球菌病 (LOD,出生后 7 至 89 天)。此外,还有 9 万名儿童死亡,其中近一半发生在撒哈拉以南非洲。2020 年,约有 46,000 例因宫内感染无乳链球菌而导致的死产和超过 500,000 例早产可能与无乳链球菌定植有关 (2)。母体无乳链球菌定植造成的众多负面影响意味着需要采取有效的预防方法来降低多种后果的风险。高收入国家在妊娠晚期末期对被无乳链球菌定植的孕妇进行筛查,并对那些被无乳链球菌定植的孕妇进行分娩期抗生素预防 (IAP)
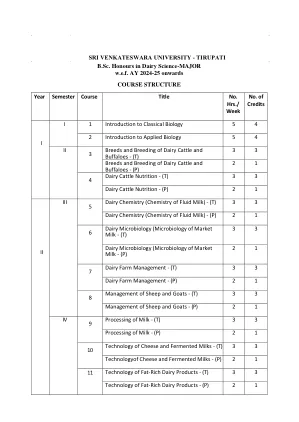
乳业科学-Tirupati
学习成果1。了解生物多样性的分类和保存原则2。了解植物的解剖,生理和生殖过程。3。关于动物分类,生理学,胚胎发展及其经济重要性的知识。4。概述细胞成分,细胞过程,例如细胞分裂,遗传和分子过程。5。理解塑造和驱动大分子和生命过程的化学原理。单元1:系统学,分类学和生态学概论1.1。系统学 - 定义和概念,分类学 - 定义和层次结构。1.2。命名法 - ICBN和ICZN,二项式和三项术命名。1.3。生态学 - 生态系统,生物多样性和保护的概念。1.4。污染和气候变化。单位2:植物学2.1的必需品。植物王国的分类。2.2。植物生理过程(光合作用,呼吸,蒸腾,植物激素)。2.3。花的结构 - 微观和宏观生成,授粉,施肥和

乳杆菌Delbrueckii Letm
L. delbrueckii le tm的delbrueckii形式具有出色的免疫调节特性。它通过非特异性和特定的联系影响先天和适应性免疫,从而根据受试者的免疫状态来控制Th1和Th2途径对免疫反应的协调。通过刺激关键细胞因子IFN,TNF,NK细胞,IL-1,IL-2和IL-6的产生来诱导特定的抗体产生并平衡人免疫系统的能力,以保护感染和癌细胞。它也是一种免疫调节剂,可以通过细胞介导的免疫来平衡和归一化的非特异性反应,并在病原剂存在下诱导更高的反应。它在慢性疾病,炎症和免疫失速(如肿瘤)中提供了免疫平衡(通过抑制)。LE菌株因此调节了对内源性和外源致病剂的免疫反应。

在酪酶基-TRNA合成酶基因YARS2相关的新生儿表型
人类遗传疾病通常是由复合杂合性突变引起的,其中突变基因的每个等位基因都具有不同的遗传病变。但是,由于缺乏适当的模型,对此类突变的研究受到阻碍。在这里,我们描述了在强制性酶二聚体中的复合异伴变体的动力学模型,该变体在一个单体中包含一个突变,而第二个单体中的另一个突变中包含一个突变。该酶由人YarS2编码用于Mito-trosyl-tRNA合成酶(MT-Tyrrs),该酶是氨基化酪氨酸到MT-TRNA Tyr的氨基酰基。yarS2是MT-氨基酰基-TRNA合成酶的基因的成员,其中致病性突变的疾病严重程度与酶活性之间的相关性有限。我们在YARS2中识别一对与新生儿死亡有关的化合物杂合变体。我们表明,虽然每个突变在MT-TYRR的同型二聚体中导致氨基酰化的最小缺陷,但反式跨性别的两个突变会协同降低酶活性,从而更大。因此,这种动力学模型准确地概括了疾病的严重程度,强调了其研究YARS2突变的效用及其对具有复合杂合突变的其他疾病的泛化潜力。

泌乳期间的孕产妇饮食干预会影响母乳和人乳微生物
1妇产科,西澳大利亚大学,珀斯,华盛顿州珀斯,澳大利亚6008; azhar.sindi@research.uwa.edu.au或asmsindi@uqu.edu.sa 2 Applied Medical Sciences学院,Umm al-Qura University,Makkah,Makkah 24381-8156,沙特阿拉伯Arabia 3 Abrastia 3 Abrastia Network,perth,Perth,WA 6000,澳大利亚,澳大利亚,澳大利亚,澳大利亚; lisa.stinson@uwa.edu.au(l.f.s. ); zoya.gridneva@uwa.edu.au(Z.G. ); donna.geddes@uwa.edu.au(d.t.g.) 4 UWA人类哺乳研究与翻译中心,珀斯,华盛顿州6009,澳大利亚5分子科学学院,西澳大利亚大学,珀斯,华盛顿州珀斯,澳大利亚6009; Mary.wlodek@uwa.edu.au或m.wlodek@unimelb.edu.au 6农业,食品和葡萄酒学校,阿德莱德大学,阿德莱德大学,阿德莱德,澳大利亚5064,澳大利亚; gabriela.leghivoyer@gmail.com(g.e.l。 ); beverly.muhlhausler@adelaide.edu.au或bev.muhlhausler@csiro.au(b.s.m.) 7妇女和儿童主题,南澳大利亚健康与医学研究所(SAHMRI),阿德莱德,SA 5000,澳大利亚; merryn.netting@adelaide.edu.au 8儿科学学科,阿德莱德大学,阿德莱大学,阿德莱德大学,SA 5006,澳大利亚9号妇女和儿童医院,阿德莱德,澳大利亚10号阿德莱德,澳大利亚10号,澳大利亚10号,妇产科,墨尔本大学,墨尔本大学,澳大利亚墨尔本大学,澳大利亚,澳大利亚3010年,澳大利亚,5000年,澳大利亚,澳大利亚,澳大利亚,澳大利亚3010年,5000年,统计,西澳大利亚大学,珀斯,华盛顿州6009,澳大利亚; Alethea.rea@murdoch.edu.au(A.R. ); michelle.trevenen@uwa.edu.au(M.L.T。)1妇产科,西澳大利亚大学,珀斯,华盛顿州珀斯,澳大利亚6008; azhar.sindi@research.uwa.edu.au或asmsindi@uqu.edu.sa 2 Applied Medical Sciences学院,Umm al-Qura University,Makkah,Makkah 24381-8156,沙特阿拉伯Arabia 3 Abrastia 3 Abrastia Network,perth,Perth,WA 6000,澳大利亚,澳大利亚,澳大利亚,澳大利亚; lisa.stinson@uwa.edu.au(l.f.s.); zoya.gridneva@uwa.edu.au(Z.G.); donna.geddes@uwa.edu.au(d.t.g.)4 UWA人类哺乳研究与翻译中心,珀斯,华盛顿州6009,澳大利亚5分子科学学院,西澳大利亚大学,珀斯,华盛顿州珀斯,澳大利亚6009; Mary.wlodek@uwa.edu.au或m.wlodek@unimelb.edu.au 6农业,食品和葡萄酒学校,阿德莱德大学,阿德莱德大学,阿德莱德,澳大利亚5064,澳大利亚; gabriela.leghivoyer@gmail.com(g.e.l。 ); beverly.muhlhausler@adelaide.edu.au或bev.muhlhausler@csiro.au(b.s.m.) 7妇女和儿童主题,南澳大利亚健康与医学研究所(SAHMRI),阿德莱德,SA 5000,澳大利亚; merryn.netting@adelaide.edu.au 8儿科学学科,阿德莱德大学,阿德莱大学,阿德莱德大学,SA 5006,澳大利亚9号妇女和儿童医院,阿德莱德,澳大利亚10号阿德莱德,澳大利亚10号,澳大利亚10号,妇产科,墨尔本大学,墨尔本大学,澳大利亚墨尔本大学,澳大利亚,澳大利亚3010年,澳大利亚,5000年,澳大利亚,澳大利亚,澳大利亚,澳大利亚3010年,5000年,统计,西澳大利亚大学,珀斯,华盛顿州6009,澳大利亚; Alethea.rea@murdoch.edu.au(A.R. ); michelle.trevenen@uwa.edu.au(M.L.T。)4 UWA人类哺乳研究与翻译中心,珀斯,华盛顿州6009,澳大利亚5分子科学学院,西澳大利亚大学,珀斯,华盛顿州珀斯,澳大利亚6009; Mary.wlodek@uwa.edu.au或m.wlodek@unimelb.edu.au 6农业,食品和葡萄酒学校,阿德莱德大学,阿德莱德大学,阿德莱德,澳大利亚5064,澳大利亚; gabriela.leghivoyer@gmail.com(g.e.l。); beverly.muhlhausler@adelaide.edu.au或bev.muhlhausler@csiro.au(b.s.m.)7妇女和儿童主题,南澳大利亚健康与医学研究所(SAHMRI),阿德莱德,SA 5000,澳大利亚; merryn.netting@adelaide.edu.au 8儿科学学科,阿德莱德大学,阿德莱大学,阿德莱德大学,SA 5006,澳大利亚9号妇女和儿童医院,阿德莱德,澳大利亚10号阿德莱德,澳大利亚10号,澳大利亚10号,妇产科,墨尔本大学,墨尔本大学,澳大利亚墨尔本大学,澳大利亚,澳大利亚3010年,澳大利亚,5000年,澳大利亚,澳大利亚,澳大利亚,澳大利亚3010年,5000年,统计,西澳大利亚大学,珀斯,华盛顿州6009,澳大利亚; Alethea.rea@murdoch.edu.au(A.R.); michelle.trevenen@uwa.edu.au(M.L.T。)13数学和统计,默多克大学,默多克,华盛顿州6150,澳大利亚 *通信:matthew.payne@uwa.edu.au.au†在澳大利亚母乳喂养 +哺乳研究与科学翻译会议上发表(澳大利亚和哺乳动物会议,2023年),澳大利亚,澳大利亚,11月2023日。