XiaoMi-AI文件搜索系统
World File Search System阴离子
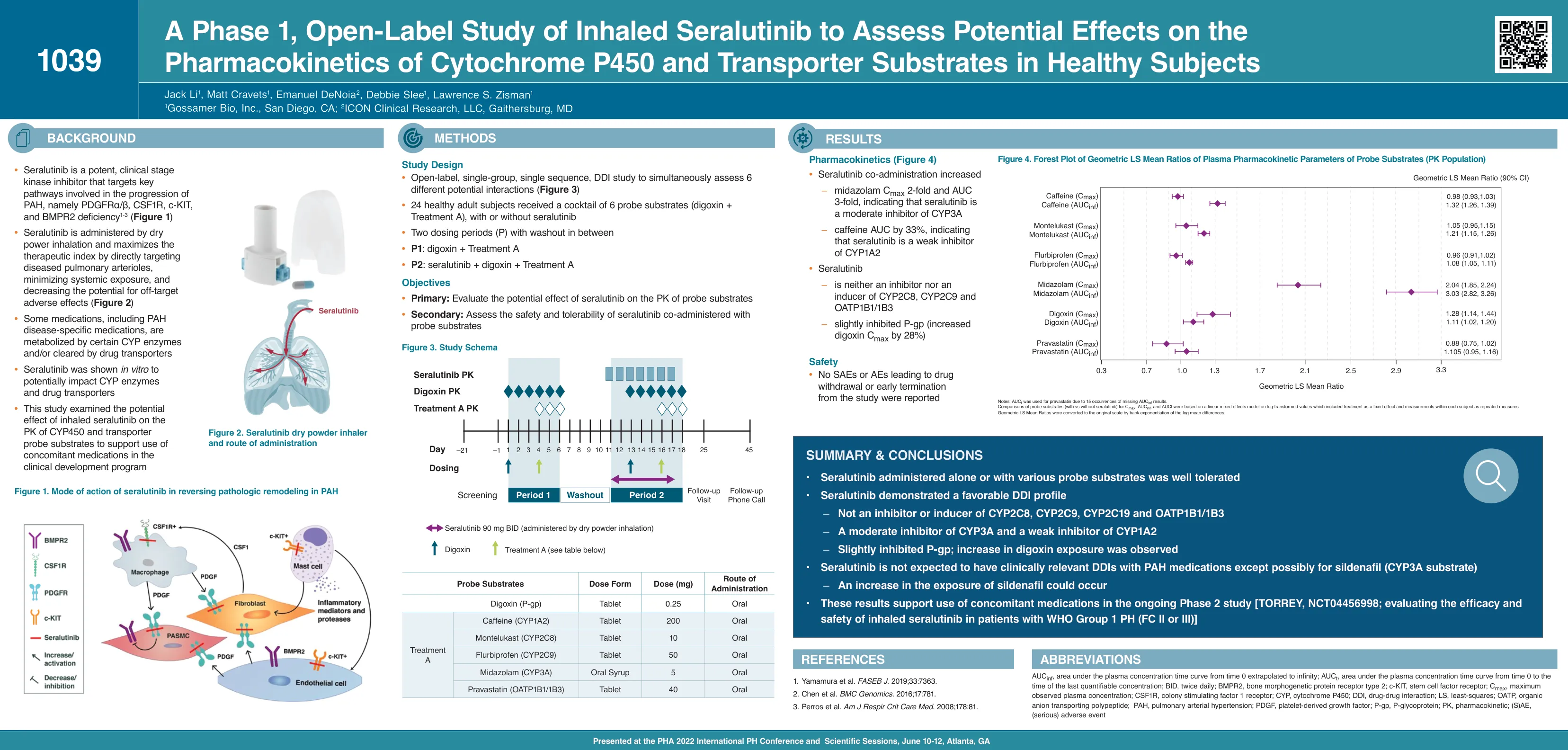
对吸入Seralutinib的1阶段的开放标签研究...
AUC INF,从推断到无穷大的时间0的等离子体浓度时间曲线下的面积;从时间0到上次可量化浓度的时间,血浆浓度时间曲线下的AUC T,面积;出价,每天两次; BMPR2,骨形态发生蛋白受体2型; C-KIT,干细胞因子受体; c最大,最大观察到的血浆浓度; CSF1R,菌落刺激因子1受体; CYP,细胞色素P450; DDI,药物相互作用; LS,最小二乘; OATP,有机阴离子运输多肽; PAH,肺动脉高压; PDGF,血小板衍生的生长因子; P-gp,p-糖蛋白; PK,药代动力学; (s)AE,(严重)不利事件

用于 RNA 递送的可电离脂质工具箱 - Mitchell 实验室
需要可离子化脂质 广义上讲,核糖核酸 (RNA) 疗法包括反义寡核苷酸 (ASO)、小干扰 RNA (siRNA)、微小 RNA (miRNA)、信使 RNA (mRNA) 和单向导 RNA (sgRNA) 介导的 CRISPR-Cas9 系统,它们可以通过不同的作用方式操纵基本上任何感兴趣的基因 1 。然而,RNA 疗法易受核酸酶影响,并且由于其体积大且带负电荷而无法渗透细胞。通过可临床转化的脂质纳米颗粒 (LNP) 将 RNA 递送至靶细胞为应对包括 COVID-19 在内的一系列危及生命的疾病提供了巨大的机会 2 。LNP 通常由四种成分组成——可离子化脂质、磷脂、胆固醇和聚乙二醇化脂质,其中可离子化脂质在保护 RNA 和促进其胞浆运输方面起主要作用。可离子化脂质在酸性 pH 下带正电荷以将 RNA 浓缩为 LNP,但在生理 pH 下呈中性以最大程度地降低毒性。它们可以在细胞摄取后在酸性内体中质子化,并与阴离子内体磷脂相互作用形成与双层膜不相容的锥形离子对(图 1)。这些阳离子-阴离子脂质对驱动从双层结构到倒六边形 H II 相的转变,从而促进膜融合/破裂、内体逃逸和货物释放到细胞溶胶 3 。自 2008 年以来,已经创建了具有多种化学特性的可离子化脂质。根据这些脂质的结构对其进行系统分类可以极大地有利于该领域并促进下一代可离子化脂质的开发。目前,有五种主要的可离子化脂质类型被广泛用于 RNA 递送(图 1)。

加工对先进费米能级的影响...
材料中的费米能通常由电荷中性决定。只要材料是纯净的并且完全符合化学计量,就可以实现电荷中性。如果添加掺杂剂或材料变为非化学计量,则会产生电荷。阴离子位点的非化学计量取决于气体气氛。样品处理过程中的阳离子非化学计量可能是由于物种的挥发性或固体溶液形成过程中的不同溶解度造成的。无论如何,目标相的合成都依赖于材料补偿其加工过程中产生的电荷的能力。例如,可以通过在价带或导带中添加电子电荷来建立电荷中性。这并不总是可能的,例如在绝缘体中。在氧化物的情况下,材料可以与周围大气交换氧气。在
二氧化硅在离子交换系统中的行为
硅酸的电离性很差。在 pH 值为中性时,水中存在的几乎所有二氧化硅都是分子而不是离子。尽管强碱树脂能够分解盐,但分子二氧化硅无法通过离子交换途径进入离子交换珠,并且受到其扩散到珠子中的速率的限制。氯化物形式的阴离子树脂去除的入口二氧化硅不到 5%,部分原因是扩散限制,部分原因是二氧化硅在 pH 值为中性时电离不利。扩散限制也是二氧化硅选择性混合物和吸附剂去除二氧化硅缓慢且不完全的主要原因。二氧化硅到达吸附位点需要很长时间,比水通常与介质接触的时间要长得多。

质膜 H+-ATPases 在矿物质营养和作物改良中的作用
质膜 H + -ATPases (PMA) 通过消耗 ATP 将 H + 从细胞质中泵出,从而产生膜电位和质子动力,以便营养物质跨膜转运进出植物细胞。PMA 通过调节根系生长、营养物质吸收和转运以及与丛枝菌根建立共生关系来参与营养物质的获取。在营养胁迫下,PMA 被激活以泵出更多的 H + 并促进有机阴离子排泄,从而提高根际营养物质的有效性。本文我们综述了 PMA 在植物有效获取和利用各种营养物质方面的生理功能和潜在分子机制的最新进展。我们还讨论了 PMA 在提高作物产量和品质方面的应用前景。


Bio-Pure人血清白蛋白(HSA),10%,20%
1产品描述和重要笔记 - 生物纯净的人血清白蛋白是一种无XENO的补充剂,尤其有资格在未分化的多能人体胚胎(HES)细胞(HES)细胞和诱导的多能茎(HIPS)细胞的生长和扩展中,均具有依赖和饲养者的无饲料和饲养者的条件。人血清白蛋白(HSA)是一种培养基补充剂,其中含有高分子量的高度溶解渗透蛋白。-HSA可有效地维持HES细胞生长和扩张。它在维持细胞膜稳定性方面特别有价值。HSA既可用于结合阴离子,阳离子和中性分子,又有其隔离和稳定多种离子和其他小分子的能力。

具有单极性驱动的可逆促进至抑制转换的电化学驻极体耦合有机突触
突触可塑性对于模仿感觉知觉、学习、记忆和遗忘具有基本意义。[1 − 3] 它通过控制突触前事件的发生来加强或削弱神经元间的连接,以突触后电流 (PSC) 为输出,从而实现对过程的定量监测。[4,5] 例如,通过重复的突触前刺激可以实现促进,从而增强超快突触传递和记忆巩固。[6] 相反,相反的过程是抑制,它代表一种抑制操作,避免过度兴奋并维持神经网络的稳定性。 [7] 由于突触可塑性在人工智能中起着促进人机交互的关键作用,人们投入了大量精力利用有机共轭材料模拟生物突触,旨在编码和放大信息。 [8 − 16] 特别是电解质门控有机材料在通道中结合了电荷传输和电化学掺杂, [17 − 19] 因此它们代表了赋予突触装置独特电性能的多功能平台。 [20 − 23] 将它们集成到光电装置中的努力导致了有机电化学晶体管 (OECT) 的发展。 [19] 作为电子突触,OECT 中离子掺杂和去掺杂的动力学已经被开发来模拟促进和抑制行为。 [10,20] 作为一种模型系统,电解质门控的 PEDOT:PSS 因可移动离子和聚合物骨架之间的可逆电化学相互作用而受到研究。[9,11] 在静电门控下,移动阴离子被驱动掺杂通道,增加通道电导率,从而产生促进作用。通过反转静电门控的极性,渗透到通道中的阴离子被提取出来,从而有可能按照抑制过程恢复到原始状态。通过掌握这种极性诱导的开关,已经实现了各种具有复杂功能的有机突触。[15] 在使用水性电解质[9,10,16]离子凝胶[14,17,23]和聚电解质门控[12]时,它们同时以电子双层 (EDL) 的形成为特征

增强用于结构研究的DNA模拟折叠剂的特征
DNA模拟折叠剂是带有阴离子侧链的芳族寡酰胺,在水中采用螺旋折叠构象,再现B- DNA双螺旋的形状和电荷表面分布。[1]它们已被证明比DNA本身更好地与某些DNA结合蛋白结合,因此是竞争性抑制蛋白-DNA相互作用的候选者。从概念上讲,DNA模拟折叠剂与天然发生的DNA模拟蛋白有关,[2]即,模仿DNA的形状和表面特征的蛋白质,并激发了非天然蛋白的设计,以干扰DNA-蛋白相互作用。[3]也可以将DNA模拟折叠剂与已提出靶向DNA结合蛋白(如转录因子)的所谓诱饵寡核苷酸进行比较。[4]

Siritanaratkul、Bhavin、Preetam Sharma、Eileen Yu 和 Alexander J. Cowan。2023 年。“提高双极电池的稳定性、选择性和电池电压
为规避碳酸盐形成问题,人们提出了各种电池配置。[5] 最有前途的配置之一是双极膜电解槽 (BPM),它由阳离子交换层 (CEL) 和阴离子交换层 (AEL) 组成。BPM 可以减轻碳酸盐的形成以及 CO 2 和产物的交叉,[6] 此外,它还允许在阴极和阳极的不同 pH 值下进行稳态操作。[7] 在正向偏置 BPM 配置中,AEL 朝向阴极,碳酸根和氢氧根离子通过 AEL 传输。[8] 虽然 AEL 朝向阴极可以提供局部碱性环境(从而使碳产品具有较高的法拉第效率),但在 CEL/AEL 界面处产生的水和/或 CO 2 可能会导致 BPM 起泡和分层。[9]