XiaoMi-AI文件搜索系统
World File Search System雄性不育

通过 ddRAD-seq 和重测序分析检测生菜中的候选基因 LsACOS5 并开发雄性不育的 InDel 标记
利用雄性不育性进行 F 1 杂交的新育种方法将为自花授粉作物莴苣育种开辟一个令人兴奋的新领域。雄性不育性是 F 1 杂交育种的一个关键性状。绘制利用雄性不育性的致病基因图谱至关重要。“CGN17397”的 ms-S 雄性不育 (MS) 基因通过 ddRAD-seq 定位到连锁群 (LG) 8,并使用两个 F 2 群体将其缩小到两个标记之间。该区域跨越约 10.16 Mb,其中根据莴苣参考基因组序列(版本 8 来自“Salinas”)注释了 94 个基因。 MS 系“CGN17397-MS”和雄性不育 (MF) 系“CGN17397-MF”的全基因组测序表明,只有一个基因在 Lsat_1_v5_gn_8_148221.1 区域有所不同,该基因是酰基辅酶 A 合成酶 5 (ACOS5) 的同源物,并且在 MS 系中被删除。据报道,ACOS5 是花粉壁形成所必需的,并且 ACOS5 的无效突变体在某些植物中完全是雄性不育的。因此,我得出结论,指定为 LsACOS5 的 Lsat_1_ v5_gn_8_148221.1 是 ms-S 基因座的生物学上合理的候选基因。利用 LsACOS5 的结构多态性,开发了 InDel 标记来选择 MS 性状。这里获得的结果为生菜的基因雄性不育提供了有价值的信息。
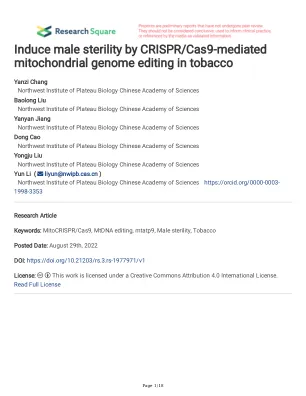
利用 CRISPR/Cas9 介导的线粒体基因组编辑诱导烟草雄性不育
农杆菌。我们再生了 76 株独立的转基因植物,并检查了单个花序的育性(补充图 1)。八株转基因植物产生了突变花。根据表型,突变花可分为两种类型:花瓣状雄蕊型(PST),具有花瓣状雄蕊和异常花药,以及败育雄蕊型(AST),花丝缩短,花药开裂异常(图 1c)。与野生型花相比,通过 TCC 染色和随后的显微镜观察,PST 和 AST 花的花粉数量较少,且活力为零(图 1d)。在 8 株发生突变花的转基因植株中,4 株(Nmu44、Nmu52、Nmu70 和 Nmu80)表现出正常花和 PST 花,2 株(Nmu43 和 Nmu46)表现出正常花和 AST 花,这 6 株均为嵌合突变体,而 2 株(Nmu58 和

阿帕里西奥·莫雷诺,拉斐尔.pdf
3.2.1 单倍体系的诱导 ...................................................................................................................................... 17 3.2.2 雄性不育系的产生 ...................................................................................................................................... 18 3.2.3 杂种优势的固定 ...................................................................................................................................... 19 3.2.4 自交不亲和性的操控 ...................................................................................................................................... 20 3.2.5 植物育种技术中的其他应用 ...................................................................................................................... 20 3.3 加速作物驯化 ............................................................................................................................................. 21

巴西和EMBRAPA的基因编辑发展
• 插入基因以诱发雄性不育(四环素) • 转基因雄性与野生雄性竞争,减少总种群数量。 • 最近,在巴西巴伊亚州茹阿泽鲁,科学家向他们展示了第一批结果,该实验是在蚊虫滋生严重的开放区域进行的。


原生质体技术和家族中的体细胞杂交
摘要:apiaceae家族的物种占据了主要市场份额,但迄今为止取决于开放的授粉品种。这导致缺乏生产统一性和降低的质量,从而促进了杂种种子的产生。困难的渗透性emanculation导致育种者使用包括体细胞杂交在内的生物技术方法。我们讨论了原生质体技术在商业特征(例如CMS(细胞质雄性雄性不育)),GMS(遗传性雄性不育)和EGMS(环境敏感的遗传无效性)等商业性状的体细胞杂种,cybrid和体外繁殖中的开发。还讨论了CMS及其候选基因的分子机制。基于摘除剂(伽马射线,X射线和紫外线)以及代谢中使用化学物质(例如碘乙酰胺或碘乙酸酯)的原生质体的饮食策略。融合原生质体的差异荧光染色通常可以用非毒性蛋白来代替新的标记方法。在这里,我们专注于初始的植物材料和组织源,用于原生质体隔离,测试的各种消化酶混合物以及对细胞壁再产生的理解,所有这些都干预了体细胞杂种再生。尽管没有躯体杂交的替代方法,但在最近的针对性状识别和选择的繁殖计划中,还讨论了各种方法,即机器人平台,人工智能,人工智能。

植物细胞器基因组育种的潜力
Maeda, A., S. Takenaka, T. Wang, B. Frink, T. Shikanai 和 M. Takenaka (2022) DYW 脱氨酶结构域对靶标 RNA 编辑位点的邻近核苷酸有明显的偏好。Plant J. 111: 756–767。Melonek, J., J. Duarte, J. Martin, L. Beuf, A. Murigneux, P. Varenne, J. Comadran, S. Specel, S. Levadoux, K. Bernath-Levin 等人 (2021) 小麦细胞质雄性不育和育性恢复的遗传基础。Nat. Commun. 12: 1036。Mok, BY, MH de Moraes, J. Zeng, DE Bosch, AV Kotrys, A. Raguram, F. Hsu, MC Radey, SB Peterson, VK Mootha 等人(2020) 细菌胞苷脱氨酶毒素可实现无 CRISPR 的线粒体碱基编辑。《自然》583:631-637。 Mok, YG, S. Hong, S.-J. Bae, S.-I. Cho 和 J.-S. Kim (2022) 植物叶绿体 DNA 的靶向 A 到 G 碱基编辑。《自然植物》8:1378-1384。 Motomura, K., Z. Moromizato 和 S. Adaniya (2003) 源自 Oryza rufipogon 的水稻品系 RT102 细胞质雄性不育的遗传和育性恢复。《日本热带农业杂志》 47: 70–76. Nakazato, I., M. Okuno, H. Yamamoto, Y. Tamura, T. Itoh, T. Shikanai, H. Takanashi, N. Tsutsumi 和 S. Arimura (2021) 拟南芥质体基因组中的靶向碱基编辑。纳特。植物 7:906–913。 Nakazato, I.、M. Okuno、C. Zhou、T. Itoh、N. Tsutsumi、M. Takenaka 和 S. Arimura (2022) 拟南芥线粒体基因组中的靶向碱基编辑。过程。国家。阿卡德。科学。美国 119:e2121177119。 Nakazato, I., M. Okuno, T. Itoh, N. Tsutsumi 和 S. Arimura (2023) 质体基因组碱基编辑器 ptpTALECD 的表征与开发。Plant J. 115: 1151–1162。Omukai, S., SI Arimura, K. Toriyama 和 T. Kazama (2021) 线粒体开放阅读框 352 的破坏可部分恢复细胞质雄性不育水稻花粉的发育。Plant Physiol. 187: 236–246。Takei, H., K. Shirasawa, K. Kuwabara, A. Toyoda, Y. Matsuzawa, S. Iioka 和 T. Ariizumi (2021) 两个番茄祖先 Solanum pimpinellifolium 和 Solanum lycopersicum var 的从头基因组组装。 cerasiforme,通过长读测序。DNA

基因组编辑技术现状及其在植物育种中的应用
H. Sugaya、A. Toyoda、T. Itoh、N. Tsutsumi 等人。 (2019)通过 TALEN 介导的线粒体基因组编辑治愈细胞质雄性不育。纳特。植物 5:722–730。 Mok, YG, S. Hong, S.-J. Bae,S.-I. Cho 和 J.-S. Kim (2022) 针对植物叶绿体 DNA 进行 A 到 G 碱基编辑。纳特。植物 8:1378–1384。 Nakazato , I. , M. Okuno , H. Yamamoto , Y. Tamura , T. Itoh , T. Shikanai , H. Takanashi , N. Tsutsumi 和 S. Arimura ( 2021 ) 拟南芥质体基因组中的靶向碱基编辑。纳特。植物 7:906–913。

CRISPR-Cas技术为创造新型玉米种质开启新时代
玉米 ( Zea mays ) 是世界上最重要的粮食作物之一,全球产量最大,为满足人类对食物、动物饲料和生物燃料的需求做出了贡献。随着人口增长和环境恶化,迫切需要采取高效、创新的育种策略来开发高产抗逆的玉米品种,以保障全球粮食安全和可持续农业。CRISPR-Cas 介导的基因组编辑技术 (CRISPR-Cas (CRISPR-associated)) 已成为植物科学和作物改良的有效而有力的工具,并且可能以不同于杂交和转基因技术的方式加速作物育种。在本综述中,我们总结了 CRISPR-Cas 技术在玉米基因功能研究和新种质生成中的应用现状和前景,以提高产量、特种玉米、植物结构、应激反应、单倍体诱导和雄性不育。本文还简要回顾了玉米基因编辑和遗传转化系统的优化。最后,讨论了使用 CRISPR-Cas 技术进行玉米遗传改良所带来的挑战和新机遇。
疫苗说明书第 13.1 节的内容
腺病毒 4 型和 7 型活疫苗口服剂尚未被评估为具有致癌或致突变潜力,或对雄性生育力的损害。 * 炭疽 AVA (BioThrax) 在一项使用怀孕兔子的发育毒性研究中评估了 BioThrax 对胚胎-胎儿和断奶前发育的影响。一组兔子在怀孕前和器官形成期(怀孕第 7 天)被施用 BioThrax 两次。第二组兔子在怀孕前和怀孕第 17 天被施用 BioThrax 两次。BioThrax 以 0.5 毫升/只兔子/次的剂量通过肌肉注射施用。未观察到对交配、生育力、怀孕、分娩、哺乳、胚胎-胎儿或断奶前发育的不良影响。本研究未发现与疫苗相关的胎儿畸形或其他致畸证据。 (CYFENDUS) CYFENDUS 尚未被评估为对动物的致癌性、致突变性或雄性不育性。给雌性大鼠服用 CYFENDUS 对生育力没有影响 * 基孔肯雅病毒