XiaoMi-AI文件搜索系统
World File Search System雄性不育

胞质雄性不育的分子基础和水稻中的生育能力 预先查看证明 使用脑电图beta/alpha 评估认知能力下降 生物。 Pharm。公牛。常规文章RSK抑制会引起... 光学量子计算机的新时代 文学
摘要细胞质男性不育(CMS)是一种母体遗传的性状,会导致花粉和花药发育中的功能障碍。cms是由核和线粒体基因组之间的相互作用引起的。通过线粒体基因组编码的引起CMS的基因的产物会影响线粒体功能和核基因的调节,从而导致雄性不育。相反,核基因组中生育基因(RF基因)的修复剂抑制了引起CMS的基因的表达并恢复男性生育力。同种质CMS系通常是由于核取代而繁殖的,这会导致去除功能性RF基因,并允许在线粒体中表达引起CMS的基因。CMS/ RF系统是理解植物中线粒体和核基因组的遗传相互作用和合作功能的绝佳模型,并且也是杂交种子生产的农艺上重要特征。在这篇评论文章中,描述了CMS,CMS相关的线粒体基因,RF基因的花粉和花药表型以及引起花粉流产的机制及其对水稻的农艺应用。

BSAseq:一种基于网络的交互式集成工作流程,用于识别大量 F2 群体中的因果突变
摘要摘要:随着新一代测序 (NGS) 技术的进步和这些技术成本的降低,批量分离分析 (BSA) 不仅成为绘制数量性状基因座 (QTL) 的有力工具,而且成为识别感兴趣表型的因果基因突变的有效方法。然而,由于存在背景突变和测序、基因分型和参考组装中的错误,通常很难区分真正的因果突变和背景突变。在本研究中,我们开发了 BSAseq 工作流程,其中包括一个自动化生物信息学分析流程,带有用于估计分离区域的概率模型和用于可视化结果的交互式 Shiny Web 应用程序。我们对雄性不育亲本系 (ms8) 进行了深度测序,以捕获批量 F2 数据中的大多数背景突变。我们将该工作流程应用于 11 个批量 F2 种群,并确定了每个种群中的真正因果突变。该工作流程直观而直接,方便没有生物信息学分析技能的用户采用。我们预计 BSAseq 将广泛应用于识别许多感兴趣的表型的因果突变。可用性:BSAseq 可在 https://www.sciapps.org/page/bsa 免费获取联系人:liya.wang@cshl.edu、ware@cshl.edu、zhanguo.xin@ars.usda.gov

通过早期开花诱导对杨树基因遏制系统进行有效评估
摘要 关键信息 早花系统 HSP:: AtFT 允许快速评估基于构建体 PsEND1:: barnase–barstar 的杨树基因遏制系统。转基因株系表现出花粉发育紊乱和不育。 摘要 通过花粉流从转基因或非本地植物物种向其可杂交的天然亲属进行垂直基因转移是一个主要问题。已经提出了基因遏制方法来减少甚至避免树种之间的基因流动。然而,由于代际时间长,评估树木的遗传遏制策略非常困难。在这种情况下,早期开花诱导可以更快地评估遗传遏制。虽然没有可靠的方法来诱导杨树的可育花,但最近开发了一种新的早花方法。在这项研究中,获得了含有基因构建体 PsEND1:: barnase–barstar 的早花杨树系。选择 PsEND1 启动子是因为它的早期表达模式、多功能性和产生与 barnase 基因融合的雄性不育植物的效率。 RT-PCR 证实了花朵中的 barnase 基因活性,花粉发育受到干扰,导致花朵不育。本研究开发的系统是研究森林树种基因控制的宝贵工具。

水稻热休克蛋白60-3B通过淀粉颗粒生物合成维持高温下的雄性生育能力
高温对水稻 (Oryza sativa) 的雄性育性有有害影响,但水稻雄配子体免受高温胁迫的机制尚不清楚。在这里,我们分离并鉴定了一种热敏感的雄性不育水稻突变体——热休克蛋白 60-3b (oshsp60-3b),它在最适温度下表现出正常的育性,但随着温度升高育性降低。高温会干扰 oshsp60-3b 花药中花粉淀粉颗粒的形成和活性氧 (ROS) 清除,导致细胞死亡和花粉败育。与突变体表型一致,OsHSP60-3B 在热休克反应中迅速上调,其蛋白质产物定位于质体。至关重要的是,OsHSP60-3B 的过表达增强了转基因植物花粉的耐热性。我们证实 OsHSP60-3B 与质体中的粉质胚乳 6 (FLO6) 相互作用,FLO6 是水稻花粉中淀粉颗粒形成的关键成分。Western blot 结果表明,高温下 oshsp60-3b 花药中的 FLO6 水平显著降低,表明当温度超过最佳条件时,OsHSP60-3B 是稳定 FLO6 所必需的。我们认为,在高温下,OsHSP60-3B 与 FLO6 相互作用,调节水稻花粉中的淀粉颗粒生物合成,并降低花药中的 ROS 水平,以确保水稻雄配子体正常发育。
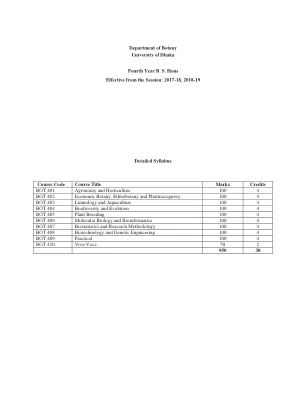
达卡大学植物学系四年级学士学位...
1. 引言:植物育种的定义、范围和目标。2. 作物的起源和驯化:作物起源中心、这一概念在植物育种中的重要性、作物的驯化。3. 植物遗传资源:种质的定义、收集、评估和保存(离体和就地),种质在植物育种计划中的应用。4. 生殖生物学和植物育种:作物的繁殖方式、植物改良中的授粉机制、自交不亲和性和雄性不育性及其在植物育种中的意义。5. 自花授粉和异花授粉作物的选择方法以及无性繁殖植物的克隆选择。6. 杂交:目标;杂交技术和类型以及人工杂交的重要性。7. 自花授粉和异花授粉作物的育种技术:大规模选择、纯系选择、谱系选择和批量方法的方法、优点和缺点。 8. 杂种优势育种:杂种优势、杂种优势和近交衰退的介绍,杂种优势的遗传基础,通过杂种优势育种取得的成果。9. 诱变育种:诱变育种介绍,植物诱变的人工诱导,诱变技术在作物改良中的应用,诱变育种的局限性。10. 回交育种:回交育种的方法、优点和局限性。11. 抗逆育种:逆境类型 - 生物和非生物,抗病育种方法,

基于同源性的候选基因的候选基因,用于高地棉花中的男性无菌性编辑(gossypium hirsutum L.)
高地棉花(Gossypium hirsutum L.)占全球棉花生产的90%以上,为全球纺织品和油料种子工业提供了天然材料。提高高地棉花产量的一种策略是增加了杂种的采用。然而,棉花的灭绝是非常耗时的,棉花雄性不育的遗传来源受到限制。在这里,我们回顾了已知的植物核男性不育(NMS)的生物化学模式,通常称为植物遗传性不育(GMS),并将其表征为四组:转录调控,剪接,脂肪酸的运输和加工以及糖的运输和加工和加工。我们已经探索了30个单子叶植物(玉米,大米和小麦)和三个双子(拟南芥,大豆和番茄)的30 gms基因的蛋白序列同源性。我们已经分析了单子植物和双子DICOT GMS基因之间的进化关系,以描述这些基因鉴定的相对相似性和相关性。五个是较低的源物种,四种是单子叶植物独有的,五核,在所有物种中有14个高度保守,而另外则有两个。使用此源,我们已经在高地棉质基因组中鉴定了23个潜在的候选基因,用于开发用于杂交棉花育种的新雄性无菌种质。将基于同源性的研究与基因组编辑结合使用可以允许发现和验证GMS基因,这些GMS基因以前在棉花中未观察到多样性,并且可能允许在杂化棉产生中使用理想的雄性无菌突变体。

CRISPR/Cas 基因组编辑在番茄改良中的应用
番茄的遗传基础狭窄,给育种带来了严峻挑战。因此,随着成簇的规律间隔短回文重复序列 (CRISPR) 相关蛋白 9 (CRISPR/Cas9) 基因组编辑的出现,快速高效的番茄育种已成为可能。番茄的许多性状已使用 CRISPR/Cas9 进行编辑和功能表征,例如植物结构和花的特性(例如叶、茎、花、雄性不育、果实、单性结实)、果实成熟、品质和营养(例如番茄红素、类胡萝卜素、GABA、TSS、花青素、保质期)、抗病性(例如 TYLCV、白粉病、晚疫病)、非生物胁迫耐受性(例如热、旱、盐度)、CN 代谢和除草剂抗性。CRISPR/Cas9 已被证明可用于将野生近缘种的优良性状从头驯化到栽培番茄,反之亦然。 CRISPR/Cas 的创新允许使用在线工具进行单向导 RNA 设计和多路复用、克隆(例如 Golden Gate 克隆、GoldenBraid 和 BioBrick 技术)、强大的 CRISPR/Cas 构建体、高效的转化方案(例如农杆菌)和用于 Cas9-gRNAs 核糖核蛋白 (RNPs) 复合物的无 DNA 原生质体方法、Cas9 变体(例如无 PAM 的 Cas12a 和 Cas9-NG/XNG-Cas9)、基于同源重组 (HR) 的双生病毒复制子基因敲入 (HKI) 以及碱基/引物编辑(Target-AID 技术)。这篇小型评论重点介绍了 CRISPR/Cas 在番茄快速高效育种方面的最新研究进展。

加速蚕豆(Vicia Faba L.)遗传增益的传统和分子育种工具
蚕豆是一种冷季豆科作物,世界各地都种植它作为食物和饲料。尽管过去蚕豆的种植面积有所减少,但由于其高种子蛋白含量和出色的生态服务功能,全球对种植蚕豆的兴趣正在增加。然而,这种作物受到各种生物和非生物胁迫,导致粮食产量不稳定、低产。虽然已经确定了对主要疾病的抗源,例如蚕豆壳针病 ( Ascochyta fabae Speg.)、锈病 ( Uromyces viciae-fabae (Pers.) Schroet.)、巧克力斑病 ( Botrytis fabae Sard.) 和瘿病 ( Physioderma viciae ),但它们的抗性只是部分的,如果没有农艺措施,就无法防止粮食产量损失。需要与宿主植物抗性基因紧密相关的 DNA 标记来增强抗性水平。在非生物胁迫方面取得的进展较少。提出了不同的育种方法,但迄今为止,基于谱系法的品系育种仍是育种计划中的主流做法。尽管如此,种子繁殖系数低、需要在防虫围栏下生长以避免杂交,以及缺乏双单倍体系统和细胞质雄性不育等工具,都阻碍了育种。这降低了育种群体的大小和育种速度,从而降低了捕获有利等位基因的稀有组合的机会。在育种计划中,蚕豆-蚕豆 (vc −) 和除草剂耐受性等 DNA 标记的可用性和使用鼓舞了育种者,并增强了对标记辅助选择的信心。与几种生物和非生物胁迫耐受性密切相关的 QTL 是可用的,它们在育种者友好平台上的验证和转换将增强选择过程。最近,基因组选择和快速育种技术以及基因组学已经触手可及,可以加速蚕豆的遗传增益。基因组资源与其他育种工具、方法和平台的进步将有助于加速育种过程,从而提高该物种的遗传增益。