XiaoMi-AI文件搜索系统
World File Search System黑素细胞

头皮巨大先天性黑素细胞痣
先天性黑素细胞痣 (CMN) 是一种表皮和真皮良性增生性皮肤病。据估计,大型至巨型 CMN 与终生恶性风险增加有关。有必要评估和监测巨型 CMN 的恶性转化风险。迄今为止,临床“ABCD”标准和免疫组织化学研究可能令人困惑,并且在某种程度上具有主观性。因此,需要阐明痣的基因组分析,以更好地了解 CMN 的恶性转化。在这里,我们描述了两个大型至巨型头皮 CMN,其潜在恶性风险的临床组织学和分子评估相反。据我们所知,这是首次对东亚大型至巨型头皮 CMN 的遗传学研究进行描述。我们建议结合仔细的病史和组织学信息来审查分子诊断,以促进对潜在恶性风险的评估。

miR-1269b 驱动人类非转移性黑素细胞对顺铂产生耐药性...
背景:据报道,miRNA 通过各种机制在多种实体肿瘤中诱导一定的耐药性。我们的研究旨在调查 miRNA-1269b 是否参与非小细胞肺癌 (NSCLC) 的化学耐药性和进展。方法:进行 MTT 和菌落形成试验以确定细胞增殖,并通过流式细胞术用膜联蛋白 V/PI 分析细胞凋亡。进行荧光素酶报告基因检测以验证 miRNA 靶向序列。在小鼠肿瘤模型中体内评估了 miR-1269b 在顺铂耐药中的作用。结果:我们发现 miR-1269b 表达在顺铂耐药的 NSCLC 标本和 NSCLC 细胞系中上调,从而促进化学耐药性和致瘤性。 miR-1269b过表达增强耐药性并促进体外细胞增殖和体内肿瘤生长,体外细胞培养中A549细胞凋亡率降低。从机制上讲,我们确定PTEN是miR-1269b的直接靶点,并且NSCLC标本中的PTEN水平与miR-1269b呈负相关。进一步研究表明,miR-1269b靶向PTEN来调节PI3K / AKT信号通路。结论:总之,这些发现表明miR-1269b / PTEN / PI3K / AKT介导的网络可能促进NSCLC对顺铂的耐药性,并且miR-1269b可以成为NSCLC患者化学耐药的潜在治疗靶点。关键词:miR-1269b、NSCLC、顺铂耐药、PTEN、PI3K/AKT 信号传导

衰老黑素细胞的信号传导过度激活头发生长
x ia at jie wy 1.2.3,#,r a l a l a l a l a l a l a l a l a l a 1.2.3,ann e Q. ph a 4,kusuk e y a s a g a g a g a g a g a a g a g a g a g a g a g a g a g a g a g a a s i i i i s ica l. f,sh,sh,sh a g 1.6,赢得了OH 7.8,shoq。 j 9. 9.10,soh ail j a d 5,chi eu n 1.2.2.11.12,trum a n k t nguy n 1.2,h和d udup 1.2,n 1.2,n 1.2,n ith shu udup sh and n ith shu udup sh and g r 1.2.13,r i z h 1,2.3,r i z h 1,2.3,k e v n H. Nobuh,nobuh至15 15,v a e s ss a M. s cfon e 2,gu a s a s a 2,k a a s a s a s a s a s a s a s a s a s a s a s a s a s a s a s a s ar,x oy是2。在n F. 1.6,Zh e,e,3.3.6.6.9,1.3.3.6.6.9,t a a a a a a a e eSco 4,a nd K. g a s a s a s a s a s a s a s a s a s a s a s s a s s a s s a s a s a s a s a s a s a s a s a s a s a s a s a s a s a s a s a s,j i 14.25,m a x m a x m a x m v。

对人工智能的看法(AI)对审计师的使用...
背景:间质干细胞衍生的外泌体(MSC-EXO)具有治疗潜力。然而,尚未阐明MSC -EXO对H 2 O 2引起的损伤后人类原代黑素细胞生存和黑色素发生的影响。因此,我们研究了MSC -EXO对人类原发性黑色素细胞的H 2 O 2生存的影响及其在体外的增殖,凋亡,衰老和黑色素发生的影响。方法:通过顺序离心从人MSC制备MSC-EXO,并以透射电子显微镜,Western blot和纳米颗粒跟踪分析为特征。人类原发性黑色素细胞分离并用不同浓度的MSC-EXO处理,然后暴露于H 2 O 2。此外,通过CCK-8,流式细胞仪,Western Blot,L-DOPA染色,酪氨酸酶活性,酪氨酸酶活性和RT-QPCR测试了使用MSC-EXO预处理对黑素细胞增殖,凋亡,衰老和黑色素生成的影响。结果:使用较低剂量的MSC -EXO保护人类原代黑素细胞免受H 2 O 2触发的凋亡的预处理,而较高剂量的MSC -EXO的预处理增强了H 2 O 2 O 2诱导的黑色素细胞细胞的细胞凋亡。Compared with the untreated control, pretreatment with a lower dose (1 µg/mL) of MSC-Exo enhanced the proliferation of melanocytes, abrogated the H 2 O 2 -increased p53, p21, IL-1β, IL-6 and IL-8 expression and partially rescued the H 2 O 2 -decreased L-dopa staining reaction, tyrosinase activity, MITF and黑素细胞中的TRP1表达。MSC-EXO可能是白癜风的有前途的治疗策略。结论:我们的发现表明,使用低剂量的MSC -EXO治疗可以通过改善H 2 O 2诱导的黑素细胞的凋亡和黑素细胞衰老来促进人类原发性黑素细胞的增殖和黑色素发生。关键词:间充质干细胞衍生的外泌体,黑素细胞,凋亡,增殖,衰老

从免疫机制揭示白癜风的治疗前景...
摘要 白癜风是一种影响全球 1% 人口的皮肤病,其特征是皮肤色素沉着丧失。它主要有两种形式:非节段性(对称性色素脱失)和节段性(局部性色素脱失)。氧化应激和黑素细胞的线粒体功能障碍会导致白癜风,而免疫特权保护毛囊黑素细胞,使色素沉着得以恢复。遗传因素和与其他自身免疫性疾病(如 1 型糖尿病和甲状腺炎)的关联表明存在可遗传的自身免疫成分。CD8+ T 细胞在白癜风中起着至关重要的作用,靶向黑素细胞并促进细胞凋亡。这些细胞与 IFN-γ 信号传导一起促进疾病进展。针对这些途径的疗法(如 JAK 抑制剂)在色素恢复方面显示出希望,尤其是与窄带 UVB 光疗法(一种金标准疗法)相结合时。外科手术,包括穿孔移植和吸水泡移植,显示出较高的效率,但会带来皮肤损伤和色素沉着的高风险。白癜风患者会遭受巨大的情感痛苦,需要心理和医学治疗。饮食干预,特别是富含抗氧化剂的饮食干预,可能有助于疾病治疗。尤其是维生素 D,它是一种很有前途的治疗剂,它通过 WNT/β-catenin 通路保护黑素细胞免受氧化应激。这篇综述指出,需要对结合免疫调节、光疗和饮食策略的靶向疗法进行更多的研究,以有效治疗白癜风。关键词:白癜风、干扰素γ、黑素细胞、Janus 激酶。

开发体内和体外模型的痤疮逆转
基础层的另一个关键成分是黑色素,它会产生黑色素,保护人体免受紫外线的侵害,并给皮肤和/或其附属物(头发,羽毛,鳞片)的色素沉着。在人类中,黑素细胞位于基础层的干细胞之间,它们通常小于基础干细胞,但具有广泛的细胞质树突,延伸到颗粒层。每平方英尺的皮肤有大约1500个黑素细胞,与基础层干细胞的比例为1:10。人们可能认为皮肤色素沉着与黑素密度有关,但实际上与黑素细胞活性有关4,5。黑色素细胞的主要作用是黑色素或黑色素发生的产生,黑色素生成是eumelanin是人类中最丰富成员的色素蛋白家族。黑色素发生发生在黑色素体中,黑色素细胞的高度专业细胞器具有酸性内部。它始于氨基酸酪氨酸,该氨基酸酪氨酸被黑素细胞特异性酶转化为多季季酮:酪氨酸酶6。一旦Eumelanin成熟,它就会包装在黑色素体内。它们通过黑色素细胞细胞质扩展向上传播,直到它们被颗粒层7的角质形成细胞释放并占据。

白癜风,从病理生理学到新兴治疗方法
白癜风是一种常见的慢性炎症性皮肤脱色症,全球患病率为 0.5–1% [ 1 ]。白斑的出现是由于表皮黑素细胞损失所致 [ 2 ]。虽然白癜风不会影响预期寿命,但颜色差异会严重影响患者的生活质量和心理健康,患者往往会遭受歧视和社会孤立,并且更容易患上精神疾病 [ 3,4 ]。到目前为止,可用的白癜风治疗方法仍然有限,治疗策略依赖于针对炎症和免疫反应的非特异性疗法,例如局部或全身使用类固醇或局部使用钙调磷酸酶抑制剂,两者均与紫外线 (UV) 光有关以促进黑素细胞再生。治疗方法的这种局限性凸显了改善白癜风管理的必要性。白癜风的病理生理学复杂,涉及多种组合因素。一种主要的假设是,免疫反应在针对黑素细胞区室中发挥作用,全基因组关联研究 (GWAS) 证明了白癜风与基因的关联,这些基因主要与先天性和适应性免疫反应有关 [ 5 ]。动物模型和人类研究也强调了免疫系统在白癜风中的重要作用,为开发针对这种疾病的靶向疗法打开了新的希望之门 [ 2 ]。这些疗法将针对疾病进展或复发过程中发生的免疫和表皮细胞炎症反应。因此,现在可以考虑早期积极治疗,以抑制针对黑素细胞的免疫反应后果,防止其丢失。一旦过度的免疫反应得到控制,
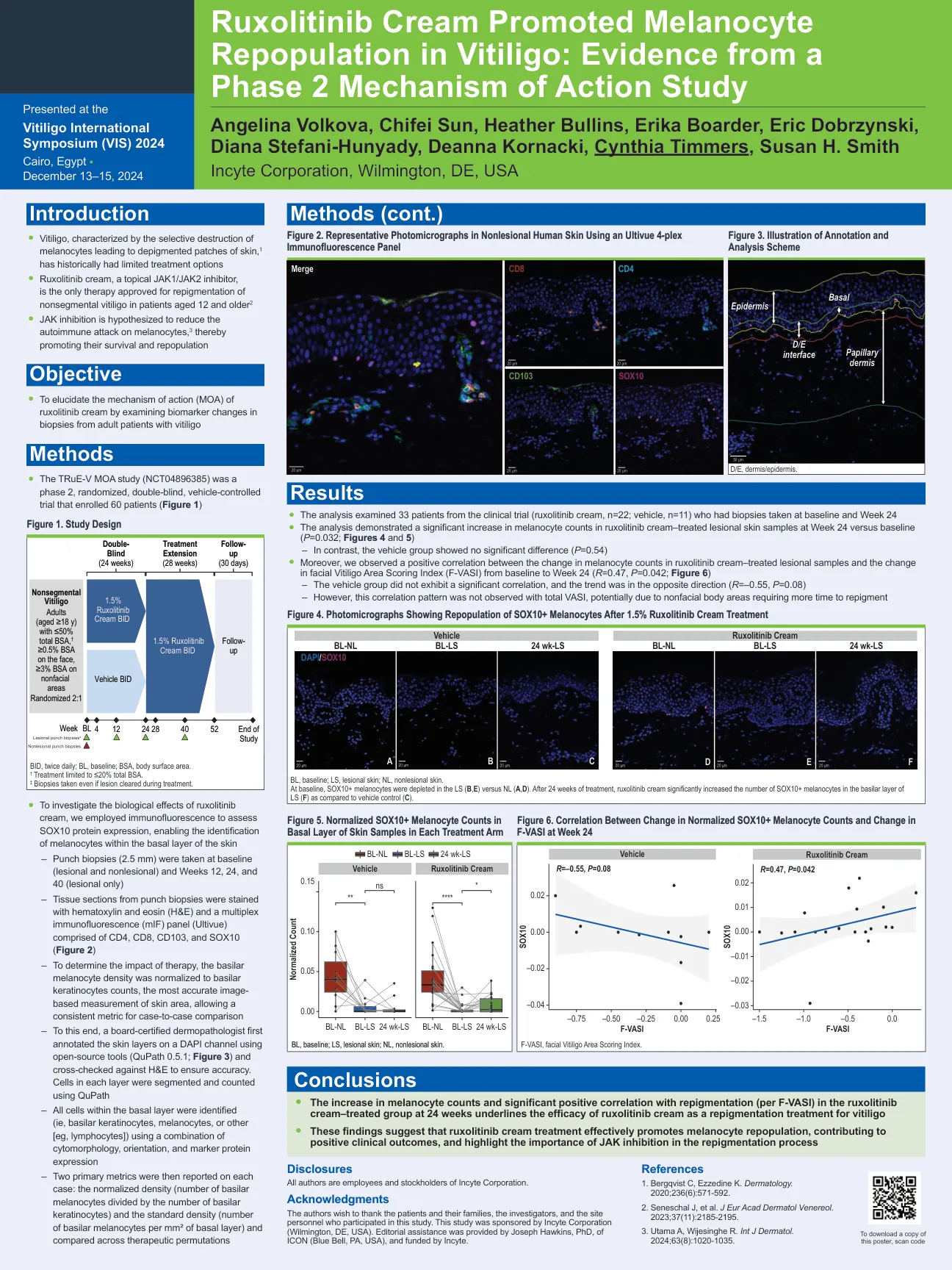
鲁索替尼乳膏促进白癜风患者黑素细胞再生:来自第 2 阶段作用机制研究的证据
BL,基线;LS,病变皮肤;NL,非病变皮肤。基线时,LS(B、E)中的 SOX10+ 黑色素细胞与 NL(A、D)相比减少。经过 24 周的治疗,与载体对照(C)相比,芦可替尼乳膏显著增加了 LS 基底层中 SOX10+ 黑色素细胞的数量(F)。

多功能的Wnt调节器是啮齿动物条纹模式的演变
信件和材料请求应发给Ricardo Mallarino。rmallarino@princeton.edu。作者贡献M.R.J.和R.M.构思了该项目并设计了实验。M.R.J. 进行了RNA-SEQ实验和大量RNA-Seq分析。 S.L. 在S.A.M.的帮助下,在条纹小鼠中进行了体外和体内基因组编辑。 和J.A.R.-P。 M.R.J. 和S.L. 对基因组编辑的动物进行了所有下游加工和分析。 下午和S.Y.S. 进行了数学建模。 C.F.G.-J. 在M.R.J.的支持下领导了SCRNA-SEQ分析。 和Q.N. M.R.J.,B.J.B. 和R.M. 进行原位杂交。 M.R.J.,B.J.B.,S.A.M。 和R.M. 进行了条纹小鼠和实验室小鼠组织的表型表征,包括免疫荧光和组织学。 M.R.J. 和S.A.M. 进行了黑素细胞细胞培养实验。 J.A.M. 进行了进化分析。 C.Y.F. 产生了横纹肌的MUS基因组和抬高注释。 J.G. 和A.P. 生成了永生的横纹纤维细胞。 M.R.J. 和R.M. 用所有作者的输入写了手稿。M.R.J.进行了RNA-SEQ实验和大量RNA-Seq分析。S.L. 在S.A.M.的帮助下,在条纹小鼠中进行了体外和体内基因组编辑。 和J.A.R.-P。 M.R.J. 和S.L. 对基因组编辑的动物进行了所有下游加工和分析。 下午和S.Y.S. 进行了数学建模。 C.F.G.-J. 在M.R.J.的支持下领导了SCRNA-SEQ分析。 和Q.N. M.R.J.,B.J.B. 和R.M. 进行原位杂交。 M.R.J.,B.J.B.,S.A.M。 和R.M. 进行了条纹小鼠和实验室小鼠组织的表型表征,包括免疫荧光和组织学。 M.R.J. 和S.A.M. 进行了黑素细胞细胞培养实验。 J.A.M. 进行了进化分析。 C.Y.F. 产生了横纹肌的MUS基因组和抬高注释。 J.G. 和A.P. 生成了永生的横纹纤维细胞。 M.R.J. 和R.M. 用所有作者的输入写了手稿。S.L.在S.A.M.的帮助下,在条纹小鼠中进行了体外和体内基因组编辑。和J.A.R.-P。 M.R.J.和S.L.对基因组编辑的动物进行了所有下游加工和分析。下午和S.Y.S. 进行了数学建模。 C.F.G.-J. 在M.R.J.的支持下领导了SCRNA-SEQ分析。 和Q.N. M.R.J.,B.J.B. 和R.M. 进行原位杂交。 M.R.J.,B.J.B.,S.A.M。 和R.M. 进行了条纹小鼠和实验室小鼠组织的表型表征,包括免疫荧光和组织学。 M.R.J. 和S.A.M. 进行了黑素细胞细胞培养实验。 J.A.M. 进行了进化分析。 C.Y.F. 产生了横纹肌的MUS基因组和抬高注释。 J.G. 和A.P. 生成了永生的横纹纤维细胞。 M.R.J. 和R.M. 用所有作者的输入写了手稿。下午和S.Y.S.进行了数学建模。C.F.G.-J. 在M.R.J.的支持下领导了SCRNA-SEQ分析。 和Q.N. M.R.J.,B.J.B. 和R.M. 进行原位杂交。 M.R.J.,B.J.B.,S.A.M。 和R.M. 进行了条纹小鼠和实验室小鼠组织的表型表征,包括免疫荧光和组织学。 M.R.J. 和S.A.M. 进行了黑素细胞细胞培养实验。 J.A.M. 进行了进化分析。 C.Y.F. 产生了横纹肌的MUS基因组和抬高注释。 J.G. 和A.P. 生成了永生的横纹纤维细胞。 M.R.J. 和R.M. 用所有作者的输入写了手稿。C.F.G.-J.在M.R.J.的支持下领导了SCRNA-SEQ分析。和Q.N.M.R.J.,B.J.B. 和R.M. 进行原位杂交。 M.R.J.,B.J.B.,S.A.M。 和R.M. 进行了条纹小鼠和实验室小鼠组织的表型表征,包括免疫荧光和组织学。 M.R.J. 和S.A.M. 进行了黑素细胞细胞培养实验。 J.A.M. 进行了进化分析。 C.Y.F. 产生了横纹肌的MUS基因组和抬高注释。 J.G. 和A.P. 生成了永生的横纹纤维细胞。 M.R.J. 和R.M. 用所有作者的输入写了手稿。M.R.J.,B.J.B.和R.M.进行原位杂交。M.R.J.,B.J.B.,S.A.M。 和R.M. 进行了条纹小鼠和实验室小鼠组织的表型表征,包括免疫荧光和组织学。 M.R.J. 和S.A.M. 进行了黑素细胞细胞培养实验。 J.A.M. 进行了进化分析。 C.Y.F. 产生了横纹肌的MUS基因组和抬高注释。 J.G. 和A.P. 生成了永生的横纹纤维细胞。 M.R.J. 和R.M. 用所有作者的输入写了手稿。M.R.J.,B.J.B.,S.A.M。和R.M.进行了条纹小鼠和实验室小鼠组织的表型表征,包括免疫荧光和组织学。M.R.J. 和S.A.M. 进行了黑素细胞细胞培养实验。 J.A.M. 进行了进化分析。 C.Y.F. 产生了横纹肌的MUS基因组和抬高注释。 J.G. 和A.P. 生成了永生的横纹纤维细胞。 M.R.J. 和R.M. 用所有作者的输入写了手稿。M.R.J.和S.A.M.进行了黑素细胞细胞培养实验。J.A.M. 进行了进化分析。 C.Y.F. 产生了横纹肌的MUS基因组和抬高注释。 J.G. 和A.P. 生成了永生的横纹纤维细胞。 M.R.J. 和R.M. 用所有作者的输入写了手稿。J.A.M.进行了进化分析。C.Y.F. 产生了横纹肌的MUS基因组和抬高注释。 J.G. 和A.P. 生成了永生的横纹纤维细胞。 M.R.J. 和R.M. 用所有作者的输入写了手稿。C.Y.F.产生了横纹肌的MUS基因组和抬高注释。J.G. 和A.P. 生成了永生的横纹纤维细胞。 M.R.J. 和R.M. 用所有作者的输入写了手稿。J.G.和A.P.生成了永生的横纹纤维细胞。M.R.J. 和R.M. 用所有作者的输入写了手稿。M.R.J.和R.M.用所有作者的输入写了手稿。
神经rest干细胞发育中的新发现
神经rest细胞是引人入胜的干细胞,通过胚胎迁移,定植各种器官并产生许多衍生物。最近,发现涉及激酶和代谢调节剂LKB1的信号传导途径在几个神经rest细胞谱系中起着至关重要的作用。这些包括皮肤中的黑素细胞,