XiaoMi-AI文件搜索系统
World File Search System切除修复

将 NEIL1 DNA 糖基化酶人工靶向至线粒体
我们专注于 DNA 修复如何帮助预防由 DNA 损伤引起的有害突变和疾病,包括线粒体 DNA (mtDNA),由于线粒体 DNA 靠近呼吸过程中产生的活性氧 (ROS),因此更容易受到损伤。我们的研究旨在增强 NEIL1 DNA 糖基化酶的线粒体定位,该酶是核和线粒体 DNA 碱基切除修复 (BER) 的关键酶。为了解决 NEIL1 内源性表达低和亚细胞定位信息有限的挑战,我们采用了一个计算框架来优化其线粒体靶向信号 (MTS),使我们能够通过共聚焦显微镜观察线粒体中的 NEIL1。通过设计一种名为 339-MTS 的新型 MTS,我们成功地增强了 NEIL1 的线粒体定位,同时保持了其在核中的存在。

引用本文:Chiara Milanese & Pier G. Mastroberardino (2020): DNA 损伤诱导的磷酸戊糖顺式通道增强作用的观点
在最近的一项研究中,我们描述了发生在小鼠模型和转录偶联和全球基因组核苷酸切除修复受损(分别为 TC-NER 和 GG-NER)患者标本中的代谢重排。在这里,我们描述了一种机制,将 DNA 修复缺陷导致的转录停滞与细胞内 ATP 水平增强联系起来,后者反过来变构抑制糖酵解酶 ATP 依赖性 6-磷酸果糖激酶(Pfk,最为人所知的是磷酸果糖激酶)通过戊糖磷酸途径(PPP)重新路由葡萄糖。PPP 的增强本质上与 NADPH 还原当量的产生增加有关——这些还原当量是在途径的氧化分支中产生的——在我们的实验系统中,氧化剂种类和/或内源性氧化还原酶活性的比例并不相符,因此最终导致还原应激 1(图 1A)。

PARP 抑制剂用于治疗小细胞肺癌及其与现有治疗方法整合的潜力
PARP 是一个蛋白质家族,它协调各种细胞过程,在 DNA 修复和基因组完整性方面发挥着重要作用。PARP1 可激活碱基切除修复 (BER),以响应 DNA 单链断裂 (SSB),其中 PARP1 与 SSB 结合并促进 DNA 修复蛋白的募集。当 PARP1 功能受损时,BER 过程会停止,并且由于复制叉不稳定而导致双链断裂 (DSB) 发生 (18)。因此,缺乏同源重组 (HR) DSB 修复途径的恶性肿瘤容易受到 PARP 抑制。PARPi 首次被证明对 BRCA1/2 突变的卵巢癌有效,而这些卵巢癌缺乏 HR (19)。随后,PARPi 的临床疗效扩展到其他携带 BRCA1/2 突变的组织学(19-27),其中大多数 PARPi 获得 FDA 批准用于治疗 BRCA1/2 突变的卵巢癌和乳腺癌(表 1)(30-37)。

解读幽门螺杆菌感染相关的线粒体 DNA 损伤和修复
线粒体基因组稳定性对于预防各种人类炎症疾病至关重要。细菌感染显著增加氧化应激,导致线粒体基因组不稳定并引发人类炎症疾病。氧化性 DNA 碱基损伤主要由细胞核 (nBER) 和线粒体 (mtBER) 中的碱基切除修复 (BER) 修复。在这篇综述中,我们总结了自发性和幽门螺杆菌感染相关的氧化性线粒体 DNA 损伤、线粒体 DNA 复制应激的分子机制及其对先天免疫信号的影响。此外,我们还讨论了位于 BER 基因的线粒体靶向序列 (MTS) 上的突变如何导致线粒体 DNA 基因组不稳定和先天免疫信号激活。总体而言,该综述总结了证据以了解线粒体基因组的动态以及 mtBER 对幽门螺杆菌相关病理结果期间先天免疫反应的影响。

DNA极性对其电子激发态放松的影响
1。Alberts,b。约翰逊(Johnson)刘易斯(J。);拉夫(M。)罗伯茨,K。 Walter,P。DNA的结构和功能。 在细胞的分子生物学中,第四版。 ;加兰科学:纽约,2002年。 2。 Hazel,P。; Huppert,J。; Balasubramanian,S。; Neidle,S。循环长度依赖性g-四链体的折叠。 J. am。 化学。 Soc。 2004,126,16405-16415。 3。 Bansal,A。; Prasad,M。;罗伊(Roy) Kukreti,S。人类甘露糖受体基因编码区的短含GC的短壁画显示出构象开关。 生物聚合物2012,97,950-962。 4。 sket,p。; Korbar,T。; Plavec,J。 D(TGGGGT)内极性位点反转的3'-3'反转对四重奏间阳离子结合的影响。 J. Mol。 结构。 2014,1075,49-52。 5。 Gupta,R。C。; Golub,E。I。; Wold,M。S。; Radding,C。M.由RECA家族的重组蛋白促进的DNA链交换的极性。 proc。 natl。 Acad.Sci。 U.S.A. 1998,95,9843-9848。 6。DeLaat,W。L。; Appeldoorn,E。; Sugasawa,K。; n。 Jaspers,N。G. J.; Hoeijmakers,J。H. J. J.人类复制蛋白A的DNA结合极性在核苷酸切除修复中核酸酶位置。 基因开发。 1998,12,2598-2609。 7。 Balasingham,S。V。; Zegeye,E。D。; H. Homberset; Rossi,M。L。; Laerdahl,J.K。; Bohr,V。A。; Tonjum,T。结核分枝杆菌DNA解旋酶XPB的酶活性和DNA底物特异性。 PLOS ONE 2012,7。 8。 nucl。Alberts,b。约翰逊(Johnson)刘易斯(J。);拉夫(M。)罗伯茨,K。 Walter,P。DNA的结构和功能。在细胞的分子生物学中,第四版。;加兰科学:纽约,2002年。2。Hazel,P。; Huppert,J。; Balasubramanian,S。; Neidle,S。循环长度依赖性g-四链体的折叠。J.am。化学。Soc。2004,126,16405-16415。 3。 Bansal,A。; Prasad,M。;罗伊(Roy) Kukreti,S。人类甘露糖受体基因编码区的短含GC的短壁画显示出构象开关。 生物聚合物2012,97,950-962。 4。 sket,p。; Korbar,T。; Plavec,J。 D(TGGGGT)内极性位点反转的3'-3'反转对四重奏间阳离子结合的影响。 J. Mol。 结构。 2014,1075,49-52。 5。 Gupta,R。C。; Golub,E。I。; Wold,M。S。; Radding,C。M.由RECA家族的重组蛋白促进的DNA链交换的极性。 proc。 natl。 Acad.Sci。 U.S.A. 1998,95,9843-9848。 6。DeLaat,W。L。; Appeldoorn,E。; Sugasawa,K。; n。 Jaspers,N。G. J.; Hoeijmakers,J。H. J. J.人类复制蛋白A的DNA结合极性在核苷酸切除修复中核酸酶位置。 基因开发。 1998,12,2598-2609。 7。 Balasingham,S。V。; Zegeye,E。D。; H. Homberset; Rossi,M。L。; Laerdahl,J.K。; Bohr,V。A。; Tonjum,T。结核分枝杆菌DNA解旋酶XPB的酶活性和DNA底物特异性。 PLOS ONE 2012,7。 8。 nucl。2004,126,16405-16415。3。Bansal,A。; Prasad,M。;罗伊(Roy) Kukreti,S。人类甘露糖受体基因编码区的短含GC的短壁画显示出构象开关。生物聚合物2012,97,950-962。4。sket,p。; Korbar,T。; Plavec,J。D(TGGGGT)内极性位点反转的3'-3'反转对四重奏间阳离子结合的影响。J. Mol。 结构。 2014,1075,49-52。 5。 Gupta,R。C。; Golub,E。I。; Wold,M。S。; Radding,C。M.由RECA家族的重组蛋白促进的DNA链交换的极性。 proc。 natl。 Acad.Sci。 U.S.A. 1998,95,9843-9848。 6。DeLaat,W。L。; Appeldoorn,E。; Sugasawa,K。; n。 Jaspers,N。G. J.; Hoeijmakers,J。H. J. J.人类复制蛋白A的DNA结合极性在核苷酸切除修复中核酸酶位置。 基因开发。 1998,12,2598-2609。 7。 Balasingham,S。V。; Zegeye,E。D。; H. Homberset; Rossi,M。L。; Laerdahl,J.K。; Bohr,V。A。; Tonjum,T。结核分枝杆菌DNA解旋酶XPB的酶活性和DNA底物特异性。 PLOS ONE 2012,7。 8。 nucl。J. Mol。结构。2014,1075,49-52。5。Gupta,R。C。; Golub,E。I。; Wold,M。S。; Radding,C。M.由RECA家族的重组蛋白促进的DNA链交换的极性。 proc。 natl。 Acad.Sci。 U.S.A. 1998,95,9843-9848。 6。DeLaat,W。L。; Appeldoorn,E。; Sugasawa,K。; n。 Jaspers,N。G. J.; Hoeijmakers,J。H. J. J.人类复制蛋白A的DNA结合极性在核苷酸切除修复中核酸酶位置。 基因开发。 1998,12,2598-2609。 7。 Balasingham,S。V。; Zegeye,E。D。; H. Homberset; Rossi,M。L。; Laerdahl,J.K。; Bohr,V。A。; Tonjum,T。结核分枝杆菌DNA解旋酶XPB的酶活性和DNA底物特异性。 PLOS ONE 2012,7。 8。 nucl。Gupta,R。C。; Golub,E。I。; Wold,M。S。; Radding,C。M.由RECA家族的重组蛋白促进的DNA链交换的极性。proc。natl。Acad.Sci。 U.S.A. 1998,95,9843-9848。 6。DeLaat,W。L。; Appeldoorn,E。; Sugasawa,K。; n。 Jaspers,N。G. J.; Hoeijmakers,J。H. J. J.人类复制蛋白A的DNA结合极性在核苷酸切除修复中核酸酶位置。 基因开发。 1998,12,2598-2609。 7。 Balasingham,S。V。; Zegeye,E。D。; H. Homberset; Rossi,M。L。; Laerdahl,J.K。; Bohr,V。A。; Tonjum,T。结核分枝杆菌DNA解旋酶XPB的酶活性和DNA底物特异性。 PLOS ONE 2012,7。 8。 nucl。Acad.Sci。U.S.A. 1998,95,9843-9848。 6。DeLaat,W。L。; Appeldoorn,E。; Sugasawa,K。; n。 Jaspers,N。G. J.; Hoeijmakers,J。H. J. J.人类复制蛋白A的DNA结合极性在核苷酸切除修复中核酸酶位置。 基因开发。 1998,12,2598-2609。 7。 Balasingham,S。V。; Zegeye,E。D。; H. Homberset; Rossi,M。L。; Laerdahl,J.K。; Bohr,V。A。; Tonjum,T。结核分枝杆菌DNA解旋酶XPB的酶活性和DNA底物特异性。 PLOS ONE 2012,7。 8。 nucl。U.S.A. 1998,95,9843-9848。6。DeLaat,W。L。; Appeldoorn,E。; Sugasawa,K。; n。 Jaspers,N。G. J.; Hoeijmakers,J。H. J. J.人类复制蛋白A的DNA结合极性在核苷酸切除修复中核酸酶位置。 基因开发。 1998,12,2598-2609。 7。 Balasingham,S。V。; Zegeye,E。D。; H. Homberset; Rossi,M。L。; Laerdahl,J.K。; Bohr,V。A。; Tonjum,T。结核分枝杆菌DNA解旋酶XPB的酶活性和DNA底物特异性。 PLOS ONE 2012,7。 8。 nucl。6。DeLaat,W。L。; Appeldoorn,E。; Sugasawa,K。; n。 Jaspers,N。G. J.; Hoeijmakers,J。H. J. J.人类复制蛋白A的DNA结合极性在核苷酸切除修复中核酸酶位置。基因开发。1998,12,2598-2609。 7。 Balasingham,S。V。; Zegeye,E。D。; H. Homberset; Rossi,M。L。; Laerdahl,J.K。; Bohr,V。A。; Tonjum,T。结核分枝杆菌DNA解旋酶XPB的酶活性和DNA底物特异性。 PLOS ONE 2012,7。 8。 nucl。1998,12,2598-2609。7。Balasingham,S。V。; Zegeye,E。D。; H. Homberset; Rossi,M。L。; Laerdahl,J.K。; Bohr,V。A。; Tonjum,T。结核分枝杆菌DNA解旋酶XPB的酶活性和DNA底物特异性。 PLOS ONE 2012,7。 8。 nucl。Balasingham,S。V。; Zegeye,E。D。; H. Homberset; Rossi,M。L。; Laerdahl,J.K。; Bohr,V。A。; Tonjum,T。结核分枝杆菌DNA解旋酶XPB的酶活性和DNA底物特异性。PLOS ONE 2012,7。8。nucl。lin,Y。H。; Chu,C.C。; Fan,H。F。; Wang,P。Y。; Cox,M。M。; Li,H。W.在没有ATP水解的情况下,5到3链交换极性是RECA核蛋白丝的内在性。ac。res。2019,47,5126-5140。9。saito,i。;高山Sugiyama,H。; Nakatani,K。通过电子传递通过电子传递进行了光诱导的DNA裂解 - 表明位于5'鸟嘌呤的鸟嘌呤残基是最含电子的位点。J.am。化学。Soc。1995,117,6406-6407。
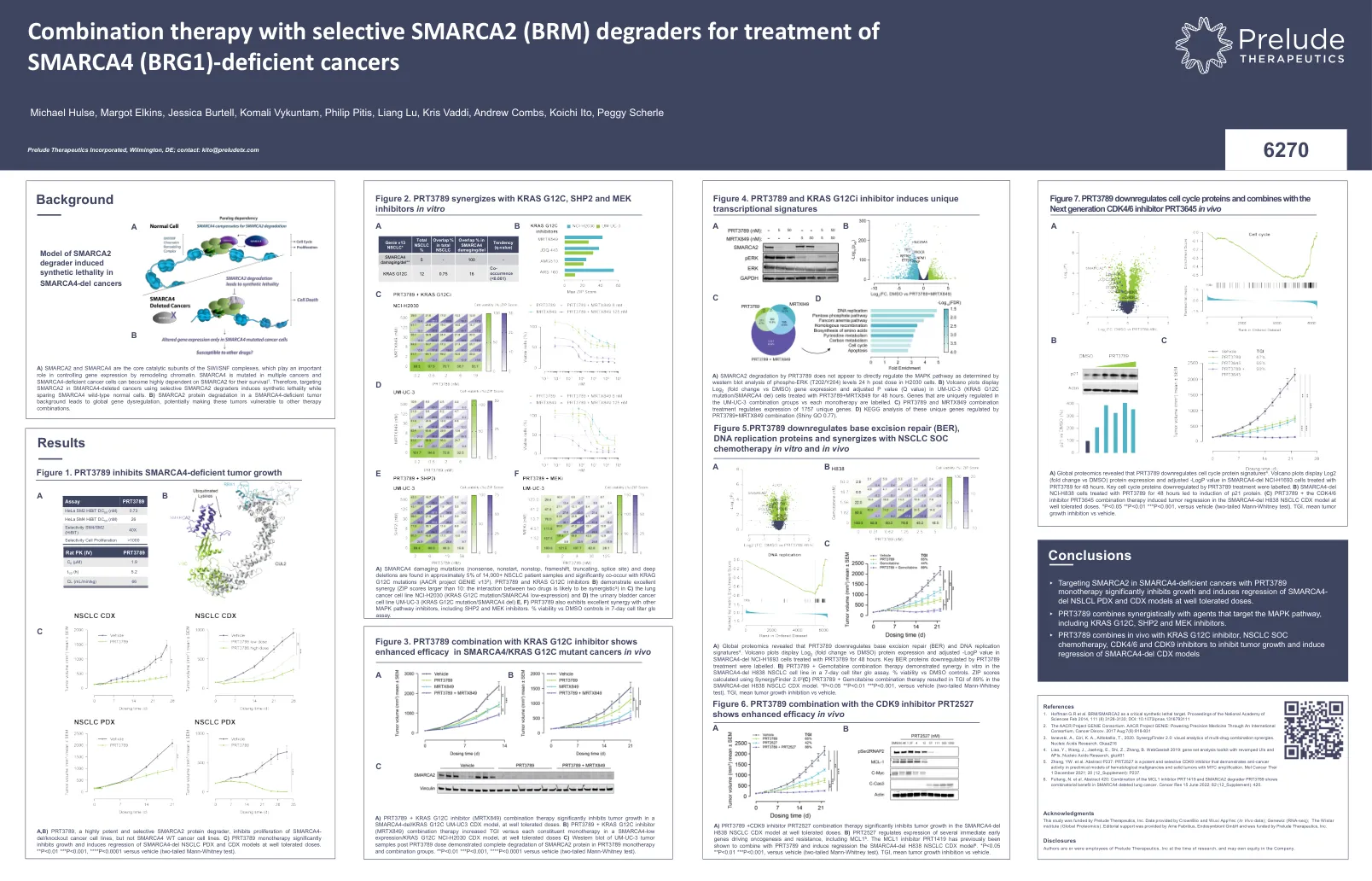
不足的癌症
a)全球蛋白质组学表明,PRT3789下调了基础切除修复(BER)和DNA复制特征4。火山图显示log 2(折叠更改与DMSO)蛋白表达和调整后的-LOGP值在Smarca4-Del NCI-H1693细胞中用PRT3789处理48小时。由PRT3789处理下调的关键BER蛋白被标记。b)PRT3789 +吉西他滨联合疗法在7天细胞滴度GLO分析中显示了SMARCA4-DEL H838 NSCLC细胞系的体外协同体外。%的生存能力与DMSO控件。使用SynergyFinder 2.0 2(C)PRT3789 +吉西他滨组合疗法计算得出的拉链得分,在SMARCA4-DEL H838 NSCLC CDX模型中,TGI为89%。*P <0.05 ** P <0.01 *** P <0.001,与车辆(两尾Mann-Whitney测试)。TGI,平均肿瘤生长抑制与车辆。

通过粗粒
蛋白XPA在核苷酸切除修复途径中起关键作用。最近的实验工作表明,XPA的功能动力学涉及沿DNA的一维扩散以搜索损伤位点。在这里,我们使用各种盐浓度的广泛的粗粒分子模拟来研究所涉及的动力学过程。结果表明扩散机制的盐浓度依赖性很强。在低盐浓度下,与旋转耦合的一维扩散是主要机制。在高盐浓度下,三维机制的扩散变得更有可能。在较广泛的盐浓度下,涉及DNA结合的残基是相似的,并且沿DNA显示的XPA的一维扩散是降低功能。此亚延伸功能暂定归因于XPA – DNA相互作用的各种强度。另外,我们表明,与DNA的结合和盐浓度升高倾向于拉伸XPA的构象,从而增加了位点的暴露范围,以结合其他修复蛋白。

下一代防辐射太空服
缩写 定义 3D 三维 ABS 丙烯腈-丁二烯-苯乙烯 AC 交流电 ALARA 尽可能低的合理值 AMF 增材制造设施 ARS 急性辐射综合症 BER 碱基切除修复 CME 日冕物质抛射 CNT 碳纳米管 CRS 慢性辐射综合症 DAP 剂量面积乘积 DAPI 4',6-二氨基-2-苯基吲哚 DC 直流电 DEP 介电泳 DMEM 杜氏改良鹰培养基 DNA 脱氧核糖核酸 DSB 双链断裂 EDTA 胰蛋白酶-乙二胺四乙酸 EMU 舱外机动装置 ESA 欧洲航天局 ESD 静电放电 EVA 舱外活动 GCR 银河宇宙辐射 Gy 格雷 HDBPE 高密度硼化聚乙烯 HDPE 高密度聚乙烯 HZE 高电荷 Z 和高能 ICRP 国际委员会放射防护 ICRU 国际辐射单位与测量委员会

E2F-1 与 Akt1 之间的潜在相互作用导致 CDDP 诱导三阴性乳腺癌细胞凋亡
针对 TNBC 的靶向治疗。2 用于治疗 TNBC 的策略之一是铂类化疗药物,例如顺式二氨二氯铂 (CDDP)。3 尽管只有一小部分 TNBC 患者对 CDDP 敏感,但由于化学耐药性导致治疗失败,其抗癌作用有限。4 人们在了解与 TNBC 化疗耐药有关的分子途径方面做出了相当大的努力。我们和其他研究小组已经表明,TNBC 细胞可以通过几种机制对 CDDP 产生耐药性,包括通过切除修复机制修复 DNA 或通过细胞周期促凋亡或抗凋亡蛋白的失调。4-7 靶向失调的细胞凋亡以缓解 TNBC 细胞的持续增殖,从而避免细胞周期进展,是一种克服化疗耐药性的有吸引力的方法。在人类癌症中,在细胞周期和细胞凋亡中发挥重要作用的两种关键蛋白质经常被激活/失调,它们是 E2F-1 和 Akt1。

DNA 修复缺陷型着色性干皮病患者皮肤癌的基因组突变图谱
着色性干皮病 (XP) 是一种由核苷酸切除修复 (NER) 途径(AG 组)或跨损伤合成 DNA 聚合酶 η (V) 基因突变引起的遗传性疾病。XP 与皮肤癌风险增加有关,对于某些群体来说,与一般人群相比,风险可高达数千倍。在这里,我们分析了来自五个 XP 组的 38 个皮肤癌基因组。我们发现 NER 的活性决定了皮肤癌基因组间突变率的异质性,并且转录偶联的 NER 超越了基因边界,降低了基因间突变率。XP-V 肿瘤中的突变谱和使用 POLH 敲除细胞系的实验揭示了聚合酶 η 在无错误绕过(i)罕见的 TpG 和 TpA DNA 损伤、(ii)嘧啶二聚体中的 3' 核苷酸和(iii)TpT 光二聚体中的作用。我们的研究揭示了 XP 皮肤癌风险的遗传基础,并对减少一般人群中紫外线诱发的突变的机制提供了见解。