XiaoMi-AI文件搜索系统
World File Search System动子

一种重写γ-球蛋白启动子并将胎儿血红蛋白重新激活的主要编辑策略
1。Frangoul,H。等。exagamglogene自动赛,用于严重的镰状细胞疾病。n Engl J Med 390,1649–1662(2024)。2。忘记,B。G。胎儿血红蛋白的遗传持久性的分子基础。ann。N. Y. Acad。 SCI。 850,38–44(1998)。 3。 Wienert,B。等。 KLF1在英国HPFH中驱动胎儿血红蛋白的表达。 血液130,803–807(2017)。 4。 Wienert,B。等。 编辑基因组,以引入与胎儿球蛋白增加有关的有益天然发生的突变。 NAT COMUM 6,7085(2015)。 5。 Martyn,G。E.等。 近端启动子中的自然调节突变通过创建从头GATA1部位来提高胎儿球蛋白表达。 血液133,852–856(2019)。 6。 Martyn,G。E.等。 自然调节突变通过破坏BCL11A或ZBTB7A结合来提升胎儿球蛋白基因。 nat Genet 50,498–503(2018)。 7。 Frati,G。等。 CRISPR-CAS9治疗镰状细胞病的安全性和功效研究突出了特异性疾病的反应。 mol ther s1525-0016(24)00470–2(2024)doi:10.1016/j.ymthe.2024.07.015。 8。 Anzalone,A。V。等。 搜索和重新固定基因组编辑,无需双链断裂或供体DNA。 自然576,149–157(2019)。 9。 Coleman,M。B.等。 am。 J. Hematol。 42,186–190(1993)。 10。 Chen,P。J.等。N. Y. Acad。SCI。 850,38–44(1998)。 3。 Wienert,B。等。 KLF1在英国HPFH中驱动胎儿血红蛋白的表达。 血液130,803–807(2017)。 4。 Wienert,B。等。 编辑基因组,以引入与胎儿球蛋白增加有关的有益天然发生的突变。 NAT COMUM 6,7085(2015)。 5。 Martyn,G。E.等。 近端启动子中的自然调节突变通过创建从头GATA1部位来提高胎儿球蛋白表达。 血液133,852–856(2019)。 6。 Martyn,G。E.等。 自然调节突变通过破坏BCL11A或ZBTB7A结合来提升胎儿球蛋白基因。 nat Genet 50,498–503(2018)。 7。 Frati,G。等。 CRISPR-CAS9治疗镰状细胞病的安全性和功效研究突出了特异性疾病的反应。 mol ther s1525-0016(24)00470–2(2024)doi:10.1016/j.ymthe.2024.07.015。 8。 Anzalone,A。V。等。 搜索和重新固定基因组编辑,无需双链断裂或供体DNA。 自然576,149–157(2019)。 9。 Coleman,M。B.等。 am。 J. Hematol。 42,186–190(1993)。 10。 Chen,P。J.等。SCI。850,38–44(1998)。 3。 Wienert,B。等。 KLF1在英国HPFH中驱动胎儿血红蛋白的表达。 血液130,803–807(2017)。 4。 Wienert,B。等。 编辑基因组,以引入与胎儿球蛋白增加有关的有益天然发生的突变。 NAT COMUM 6,7085(2015)。 5。 Martyn,G。E.等。 近端启动子中的自然调节突变通过创建从头GATA1部位来提高胎儿球蛋白表达。 血液133,852–856(2019)。 6。 Martyn,G。E.等。 自然调节突变通过破坏BCL11A或ZBTB7A结合来提升胎儿球蛋白基因。 nat Genet 50,498–503(2018)。 7。 Frati,G。等。 CRISPR-CAS9治疗镰状细胞病的安全性和功效研究突出了特异性疾病的反应。 mol ther s1525-0016(24)00470–2(2024)doi:10.1016/j.ymthe.2024.07.015。 8。 Anzalone,A。V。等。 搜索和重新固定基因组编辑,无需双链断裂或供体DNA。 自然576,149–157(2019)。 9。 Coleman,M。B.等。 am。 J. Hematol。 42,186–190(1993)。 10。 Chen,P。J.等。850,38–44(1998)。3。Wienert,B。等。 KLF1在英国HPFH中驱动胎儿血红蛋白的表达。 血液130,803–807(2017)。 4。 Wienert,B。等。 编辑基因组,以引入与胎儿球蛋白增加有关的有益天然发生的突变。 NAT COMUM 6,7085(2015)。 5。 Martyn,G。E.等。 近端启动子中的自然调节突变通过创建从头GATA1部位来提高胎儿球蛋白表达。 血液133,852–856(2019)。 6。 Martyn,G。E.等。 自然调节突变通过破坏BCL11A或ZBTB7A结合来提升胎儿球蛋白基因。 nat Genet 50,498–503(2018)。 7。 Frati,G。等。 CRISPR-CAS9治疗镰状细胞病的安全性和功效研究突出了特异性疾病的反应。 mol ther s1525-0016(24)00470–2(2024)doi:10.1016/j.ymthe.2024.07.015。 8。 Anzalone,A。V。等。 搜索和重新固定基因组编辑,无需双链断裂或供体DNA。 自然576,149–157(2019)。 9。 Coleman,M。B.等。 am。 J. Hematol。 42,186–190(1993)。 10。 Chen,P。J.等。Wienert,B。等。KLF1在英国HPFH中驱动胎儿血红蛋白的表达。血液130,803–807(2017)。4。Wienert,B。等。 编辑基因组,以引入与胎儿球蛋白增加有关的有益天然发生的突变。 NAT COMUM 6,7085(2015)。 5。 Martyn,G。E.等。 近端启动子中的自然调节突变通过创建从头GATA1部位来提高胎儿球蛋白表达。 血液133,852–856(2019)。 6。 Martyn,G。E.等。 自然调节突变通过破坏BCL11A或ZBTB7A结合来提升胎儿球蛋白基因。 nat Genet 50,498–503(2018)。 7。 Frati,G。等。 CRISPR-CAS9治疗镰状细胞病的安全性和功效研究突出了特异性疾病的反应。 mol ther s1525-0016(24)00470–2(2024)doi:10.1016/j.ymthe.2024.07.015。 8。 Anzalone,A。V。等。 搜索和重新固定基因组编辑,无需双链断裂或供体DNA。 自然576,149–157(2019)。 9。 Coleman,M。B.等。 am。 J. Hematol。 42,186–190(1993)。 10。 Chen,P。J.等。Wienert,B。等。编辑基因组,以引入与胎儿球蛋白增加有关的有益天然发生的突变。NAT COMUM 6,7085(2015)。 5。 Martyn,G。E.等。 近端启动子中的自然调节突变通过创建从头GATA1部位来提高胎儿球蛋白表达。 血液133,852–856(2019)。 6。 Martyn,G。E.等。 自然调节突变通过破坏BCL11A或ZBTB7A结合来提升胎儿球蛋白基因。 nat Genet 50,498–503(2018)。 7。 Frati,G。等。 CRISPR-CAS9治疗镰状细胞病的安全性和功效研究突出了特异性疾病的反应。 mol ther s1525-0016(24)00470–2(2024)doi:10.1016/j.ymthe.2024.07.015。 8。 Anzalone,A。V。等。 搜索和重新固定基因组编辑,无需双链断裂或供体DNA。 自然576,149–157(2019)。 9。 Coleman,M。B.等。 am。 J. Hematol。 42,186–190(1993)。 10。 Chen,P。J.等。NAT COMUM 6,7085(2015)。5。Martyn,G。E.等。近端启动子中的自然调节突变通过创建从头GATA1部位来提高胎儿球蛋白表达。血液133,852–856(2019)。6。Martyn,G。E.等。自然调节突变通过破坏BCL11A或ZBTB7A结合来提升胎儿球蛋白基因。nat Genet 50,498–503(2018)。7。Frati,G。等。CRISPR-CAS9治疗镰状细胞病的安全性和功效研究突出了特异性疾病的反应。mol ther s1525-0016(24)00470–2(2024)doi:10.1016/j.ymthe.2024.07.015。8。Anzalone,A。V。等。搜索和重新固定基因组编辑,无需双链断裂或供体DNA。自然576,149–157(2019)。9。Coleman,M。B.等。am。J. Hematol。42,186–190(1993)。 10。 Chen,P。J.等。42,186–190(1993)。10。Chen,P。J.等。Chen,P。J.等。g伽玛A伽马(β+)胎儿血红蛋白的遗传持久性:g伽玛-158 c-> t在顺式中与-175 t-> c c gamma-lobin基因的突变会导致G Gama-- gamma基因的增加导致G Gama-Globobin的增加。通过操纵细胞决定因素的编辑结果来增强质量编辑系统。Cell 184,5635-5652.E29(2021)。 11。 Ravi,N。S.等。 通过CRISPR基础编辑来识别新型HPFH样突变,从而提高了胎儿血红蛋白的表达。 Elife 11,E65421(2022)。 12。 Kim,H。K.等。 预测人类细胞中主要编辑指南RNA的效率。 nat Biotechnol(2020)doi:10.1038/s41587-020-0677-y。 13。 Nelson,J。W.等。 设计的Pegrnas提高了主要的编辑效率。 NAT生物技术40,402–410(2022)。 14。 Habib,O.,Habib,G.,Hwang,G.-H。 &Bae,S。人类胚胎干细胞中主要编辑结果的全面分析。 核酸Res 50,1187–1197(2022)。 15。 Lee,J。等。 prime用真正的cas9 nickases编辑最大程度地减少了不需要的indels。 nat Commun 14,1786(2023)。 16。 Antoniou,P。等。 基础编辑介导的γ-球蛋白顺式调节元件的解剖,用于胎儿血红蛋白表达的治疗重新激活。 nat Commun 13,6618(2022)。 17。 Pavani,G。等。 通过人类造血干细胞中α-珠蛋白基因座的CRISPR/CAS9编辑对β-丘脑的抗性。 血液Adv 5,1137–1153(2021)。 18。Cell 184,5635-5652.E29(2021)。11。Ravi,N。S.等。 通过CRISPR基础编辑来识别新型HPFH样突变,从而提高了胎儿血红蛋白的表达。 Elife 11,E65421(2022)。 12。 Kim,H。K.等。 预测人类细胞中主要编辑指南RNA的效率。 nat Biotechnol(2020)doi:10.1038/s41587-020-0677-y。 13。 Nelson,J。W.等。 设计的Pegrnas提高了主要的编辑效率。 NAT生物技术40,402–410(2022)。 14。 Habib,O.,Habib,G.,Hwang,G.-H。 &Bae,S。人类胚胎干细胞中主要编辑结果的全面分析。 核酸Res 50,1187–1197(2022)。 15。 Lee,J。等。 prime用真正的cas9 nickases编辑最大程度地减少了不需要的indels。 nat Commun 14,1786(2023)。 16。 Antoniou,P。等。 基础编辑介导的γ-球蛋白顺式调节元件的解剖,用于胎儿血红蛋白表达的治疗重新激活。 nat Commun 13,6618(2022)。 17。 Pavani,G。等。 通过人类造血干细胞中α-珠蛋白基因座的CRISPR/CAS9编辑对β-丘脑的抗性。 血液Adv 5,1137–1153(2021)。 18。Ravi,N。S.等。通过CRISPR基础编辑来识别新型HPFH样突变,从而提高了胎儿血红蛋白的表达。Elife 11,E65421(2022)。12。Kim,H。K.等。 预测人类细胞中主要编辑指南RNA的效率。 nat Biotechnol(2020)doi:10.1038/s41587-020-0677-y。 13。 Nelson,J。W.等。 设计的Pegrnas提高了主要的编辑效率。 NAT生物技术40,402–410(2022)。 14。 Habib,O.,Habib,G.,Hwang,G.-H。 &Bae,S。人类胚胎干细胞中主要编辑结果的全面分析。 核酸Res 50,1187–1197(2022)。 15。 Lee,J。等。 prime用真正的cas9 nickases编辑最大程度地减少了不需要的indels。 nat Commun 14,1786(2023)。 16。 Antoniou,P。等。 基础编辑介导的γ-球蛋白顺式调节元件的解剖,用于胎儿血红蛋白表达的治疗重新激活。 nat Commun 13,6618(2022)。 17。 Pavani,G。等。 通过人类造血干细胞中α-珠蛋白基因座的CRISPR/CAS9编辑对β-丘脑的抗性。 血液Adv 5,1137–1153(2021)。 18。Kim,H。K.等。预测人类细胞中主要编辑指南RNA的效率。nat Biotechnol(2020)doi:10.1038/s41587-020-0677-y。13。Nelson,J。W.等。设计的Pegrnas提高了主要的编辑效率。NAT生物技术40,402–410(2022)。14。Habib,O.,Habib,G.,Hwang,G.-H。 &Bae,S。人类胚胎干细胞中主要编辑结果的全面分析。 核酸Res 50,1187–1197(2022)。 15。 Lee,J。等。 prime用真正的cas9 nickases编辑最大程度地减少了不需要的indels。 nat Commun 14,1786(2023)。 16。 Antoniou,P。等。 基础编辑介导的γ-球蛋白顺式调节元件的解剖,用于胎儿血红蛋白表达的治疗重新激活。 nat Commun 13,6618(2022)。 17。 Pavani,G。等。 通过人类造血干细胞中α-珠蛋白基因座的CRISPR/CAS9编辑对β-丘脑的抗性。 血液Adv 5,1137–1153(2021)。 18。Habib,O.,Habib,G.,Hwang,G.-H。 &Bae,S。人类胚胎干细胞中主要编辑结果的全面分析。核酸Res 50,1187–1197(2022)。15。Lee,J。等。 prime用真正的cas9 nickases编辑最大程度地减少了不需要的indels。 nat Commun 14,1786(2023)。 16。 Antoniou,P。等。 基础编辑介导的γ-球蛋白顺式调节元件的解剖,用于胎儿血红蛋白表达的治疗重新激活。 nat Commun 13,6618(2022)。 17。 Pavani,G。等。 通过人类造血干细胞中α-珠蛋白基因座的CRISPR/CAS9编辑对β-丘脑的抗性。 血液Adv 5,1137–1153(2021)。 18。Lee,J。等。prime用真正的cas9 nickases编辑最大程度地减少了不需要的indels。nat Commun 14,1786(2023)。16。Antoniou,P。等。基础编辑介导的γ-球蛋白顺式调节元件的解剖,用于胎儿血红蛋白表达的治疗重新激活。nat Commun 13,6618(2022)。17。Pavani,G。等。通过人类造血干细胞中α-珠蛋白基因座的CRISPR/CAS9编辑对β-丘脑的抗性。血液Adv 5,1137–1153(2021)。18。Everette,K。A.等。在体内造血干细胞的体内质量编辑促进小鼠植入后镰状细胞疾病表型。nat Biomed Eng 7,616–628(2023)。19。Peterka,M。等。利用DSB修复以促进有效的同源性依赖性和 - 独立的质量编辑。nat Commun 13,1240(2022)。20。Magnani,A。等。对镰状细胞疾病的同种异体移植后混合嵌合体患者进行了广泛的多系数分析:对基因治疗的造血和植入阈值的见解。Haematologica 105,1240–1247(2020)。21。Sun,Y。等。 在小鼠中耐用基因校正的肺部干细胞的体内编辑。 科学384,1196–1202(2024)。 22。 Doman,J。L.等。 噬菌体辅助进化和蛋白质工程产生紧凑,有效的主要编辑者。 单元格186,3983-4002.E26(2023)。 23。 Wimberger,S。等。 同时抑制DNA-PK和POLθ提高了基因组编辑的整合效率和精度。 nat Commun 14,4761(2023)。 24。 Yan,J。等。 用内源性的小RNA结合蛋白改善原始编辑。 自然628,639–647(2024)。 25。 Levesque,S.,Cosentino,A.,Verma,A.,Genovese,P。&Bauer,D。E.通过调节核苷酸代谢,增强造血干和祖细胞中的质量编辑。 nat Biotechnol(2024)doi:10.1038/s41587-024-02266-4。 26。 核酸res。Sun,Y。等。在小鼠中耐用基因校正的肺部干细胞的体内编辑。 科学384,1196–1202(2024)。 22。 Doman,J。L.等。 噬菌体辅助进化和蛋白质工程产生紧凑,有效的主要编辑者。 单元格186,3983-4002.E26(2023)。 23。 Wimberger,S。等。 同时抑制DNA-PK和POLθ提高了基因组编辑的整合效率和精度。 nat Commun 14,4761(2023)。 24。 Yan,J。等。 用内源性的小RNA结合蛋白改善原始编辑。 自然628,639–647(2024)。 25。 Levesque,S.,Cosentino,A.,Verma,A.,Genovese,P。&Bauer,D。E.通过调节核苷酸代谢,增强造血干和祖细胞中的质量编辑。 nat Biotechnol(2024)doi:10.1038/s41587-024-02266-4。 26。 核酸res。在小鼠中耐用基因校正的肺部干细胞的体内编辑。科学384,1196–1202(2024)。22。Doman,J。L.等。噬菌体辅助进化和蛋白质工程产生紧凑,有效的主要编辑者。单元格186,3983-4002.E26(2023)。23。Wimberger,S。等。同时抑制DNA-PK和POLθ提高了基因组编辑的整合效率和精度。nat Commun 14,4761(2023)。24。Yan,J。等。 用内源性的小RNA结合蛋白改善原始编辑。 自然628,639–647(2024)。 25。 Levesque,S.,Cosentino,A.,Verma,A.,Genovese,P。&Bauer,D。E.通过调节核苷酸代谢,增强造血干和祖细胞中的质量编辑。 nat Biotechnol(2024)doi:10.1038/s41587-024-02266-4。 26。 核酸res。Yan,J。等。用内源性的小RNA结合蛋白改善原始编辑。自然628,639–647(2024)。25。Levesque,S.,Cosentino,A.,Verma,A.,Genovese,P。&Bauer,D。E.通过调节核苷酸代谢,增强造血干和祖细胞中的质量编辑。nat Biotechnol(2024)doi:10.1038/s41587-024-02266-4。26。核酸res。Brinkman,E。K.,Chen,T.,Amendola,M。&Van Steensel,B。通过序列痕量分解对基因组编辑的易于定量评估。42,E168(2014)。 27。 Brusson,M。等。 新型的慢病毒载体,用于结合基因添加和基因沉默策略的镰状细胞疾病基因治疗。 mol the核酸32,229–246(2023)。 28。 Gaudelli,N。M.等。 腺嘌呤基础编辑者的定向演变,活动增加和42,E168(2014)。27。Brusson,M。等。 新型的慢病毒载体,用于结合基因添加和基因沉默策略的镰状细胞疾病基因治疗。 mol the核酸32,229–246(2023)。 28。 Gaudelli,N。M.等。 腺嘌呤基础编辑者的定向演变,活动增加和Brusson,M。等。新型的慢病毒载体,用于结合基因添加和基因沉默策略的镰状细胞疾病基因治疗。mol the核酸32,229–246(2023)。28。Gaudelli,N。M.等。腺嘌呤基础编辑者的定向演变,活动增加和

1新型合成启动子调整基因的设计...
(未通过同行评审认证)是作者/资助者。保留所有权利。未经许可就不允许重复使用。此预印本版的版权持有人于2025年1月9日发布。 https://doi.org/10.1101/2025.01.09.632034 doi:Biorxiv Preprint

人类基因启动子处的 G-四链体蛋白质相互作用组
基因启动子处的 DNA-蛋白质相互作用在基因表达中起着至关重要的作用。人类细胞的启动子富含富含鸟嘌呤的序列,这些序列可以形成四链 G-四链体 (G4) 结构。G4 正在成为基因调控中一类独特的基于结构的调控元件,它们与蛋白质的相互作用对于 G4 的作用至关重要。目前,我们对 G4-蛋白质相互作用的理解主要是基于个案,没有系统信息。在这项工作中,我们使用来自 ENCODE 项目的数据检查了共识 G4 形成区 G4(+) 周围 1,183 种人类 DNA 结合蛋白(包括转录因子、组蛋白及其修饰酶)的空间占有率。我们发现 G4(+)、其近端侧和远端侧是三个主要的蛋白质结合位点。几乎所有蛋白质在这些位点上都富集或耗尽,这可能是由于竞争或位点之间的时空转换,导致不同程度的变化或持久性,在细胞/组织类型内或跨细胞/组织类型。值得注意的是,组蛋白被排除在 G4(+) 的近端之外,它们与 G4(+) 的结合分别通过乙酰化和甲基化打开和关闭。此外,远端优先富集 H3K23me2 和 H3K4me2。我们的实验还揭示了相应的 G4-蛋白质相互作用模式。总之,我们的结果表明 G4 在动态定义和协调基因启动子处的染色质结构和 DNA-蛋白质相互作用以进行转录调控方面发挥着普遍作用,而这项任务不太可能通过基于序列的 DNA 识别来完成。

人类基因启动子处的 G-四链体蛋白质相互作用组
基因启动子处的 DNA-蛋白质相互作用在基因表达中起着至关重要的作用。人类细胞的启动子富含富含鸟嘌呤的序列,这些序列可以形成四链 G-四链体 (G4) 结构。G4 正在成为基因调控中一类独特的基于结构的调控元件,它们与蛋白质的相互作用对于 G4 的作用至关重要。目前,我们对 G4-蛋白质相互作用的理解主要是基于个案,没有系统信息。在这项工作中,我们使用来自 ENCODE 项目的数据检查了共识 G4 形成区 G4(+) 周围 1,183 种人类 DNA 结合蛋白(包括转录因子、组蛋白及其修饰酶)的空间占有率。我们发现 G4(+)、其近端侧和远端侧是三个主要的蛋白质结合位点。几乎所有蛋白质在这些位点上都富集或耗尽,这可能是由于竞争或位点之间的时空转换,导致不同程度的变化或持久性,在细胞/组织类型内或跨细胞/组织类型。值得注意的是,组蛋白被排除在 G4(+) 的近端之外,它们与 G4(+) 的结合分别通过乙酰化和甲基化打开和关闭。此外,远端优先富集 H3K23me2 和 H3K4me2。我们的实验还揭示了相应的 G4-蛋白质相互作用模式。总之,我们的结果表明 G4 在动态定义和协调基因启动子处的染色质结构和 DNA-蛋白质相互作用以进行转录调控方面发挥着普遍作用,而这项任务不太可能通过基于序列的 DNA 识别来完成。

启动子的全基因组核小体分辨率图
摘要 增强子-启动子环路模型长期以来一直主导着基因调控领域,其中增强子通过物理接触激活其靶基因。然而,由于存在替代机制的证据以及缺乏系统验证(主要是由于缺乏合适的实验技术),该模型的普遍性受到了质疑。在本研究中,我们提出了一种新的基于 MNase 的邻近连接方法,称为 MChIP-C,该方法可以在基因组范围内以单核小体分辨率测量蛋白质介导的染色质相互作用。通过应用 MChIP-C 研究 K562 细胞中以 H3K4me3 启动子为中心的相互作用,我们发现与基于限制性内切酶的 C 方法相比,它具有大大提高的分辨率和灵敏度。这使我们能够将 EP300 组蛋白乙酰转移酶和 SWI/SNF 重塑复合物确定为建立和/或维持增强子-启动子相互作用的潜在候选者。最后,利用已发表的 CRISPRi 筛选数据,我们发现大多数经过功能验证的增强子确实与其同源启动子发生物理相互作用,支持增强子-启动子环路模型。

接下来会发生粘结启动子树脂...
Resin Developed Pre-Commercial Designation Year Developed Compostion Penacolite® B1A 1940's Resorcinol - Formaldehyde Penacolite® B16 1950's Resorcinol - Formaldehyde Penacolite® B17 1962 Resorcinol - Formaldehyde Penacolite® B18 1964 Resorcinol - Formaldehyde Penacolite® B18S RDL - 065 1976 RF Resin + RM441 Penacolite® B19S RDL - 095 1978 RF Resin + RM441 Penacolite® B20S*** RDL - 055 1988 R + Styrene + F Resin Penacolite® B21S *** RDL - 516 2004 R + Styrene + F Resin Penacolite® B22 1950's Phenol + Resorcinol + FPenaColite®untipro 100 ***良好的年份代码:“ Rajic” 2003 M-羟基二苯胺Penacolite®I868 C *** CRL -434 2009 RESORCINOL -MDI ADRAUCT -MDI ADRAUCT

ESTANE®ECO 12T85 钻石分散™HSDC1 -PDS dowerAdd™9063 -PDS 5键式 - 当选 - - - 无硫酸盐的色彩护理洗发水I Telophi™生物技术成分还原面部血清 专业纸申请的分散剂 pelethane®5863-87A-R1 TPU CD-0123(EU)即时散开2相喷雾 局部药物输送 热收缩管 Pearlbond™702 Exp 为什么医疗设备成功始于适当的生物相容性评估 Solthix™A460 -PDS SH-0212(in)丰富的洗发水凝胶 产品规范 2021公司报告 iSoplast®300ETP Tecophilic™HP系列TPU Carbopol 974p NF聚合物 药物 - 聚合物 - 典型 - - - ... 产品规范 estane®3DTPU M95A Carbopol Aqua SF-1 OS聚合物毒理学研究 Lanco™stat电导率启动子 Carbopol聚合物作为非水的流变修饰符... ESTANE®2355-85ABRTPU estane®58206TPU ESTANE®ZHF 80AT7 - 技术数据表 Pawel Balcerzak1 | Liliana Miinea | Elena Draganoiu lubrizol赋形剂(OTC)药品
lubrizol Advanced Materials,Inc。(“ Lubrizol”)希望您找到了提供的信息,但是您警告您,该材料(包括任何原型公式)仅用于信息目的,并且独自负责自己对信息的适当使用进行评估。在适用法律允许的最大范围内,Lubrizol不做任何陈述,担保或保证(无论是明示,暗示,法定还是其他),包括对特定目的的适销性或适用性的任何暗示保证,或任何信息的完整性,准确性或及时性。lubrizol不能保证此处参考的材料将如何与其他物质一起执行,以任何方法,条件或过程,任何设备或非实验室环境中的任何方法,条件或过程。在包含这些材料的任何产品进行商业化之前,您应该彻底测试该产品,包括产品包装的方式,以确定其性能,功效和安全性。您对您生产的任何产品的性能,功效和安全性负责。lubrizol不承担任何责任,您应承担所有使用或处理任何材料的风险和责任。所有司法管辖区都不得批准任何索赔。任何与这些产品相关的索赔的实体均负责遵守当地法律和法规。您承认并同意您正在使用此处提供的信息自负。如果您对Lubrizol提供的信息不满意,则您的独家补救措施将不使用信息。未经专利所有人许可,本文中没有任何内容作为许可,建议或诱因,以实践任何专利发明,而您的唯一责任有责任确定是否存在与专利侵犯与所提供信息有关的任何组件的专利侵犯或组合组合有关的问题。
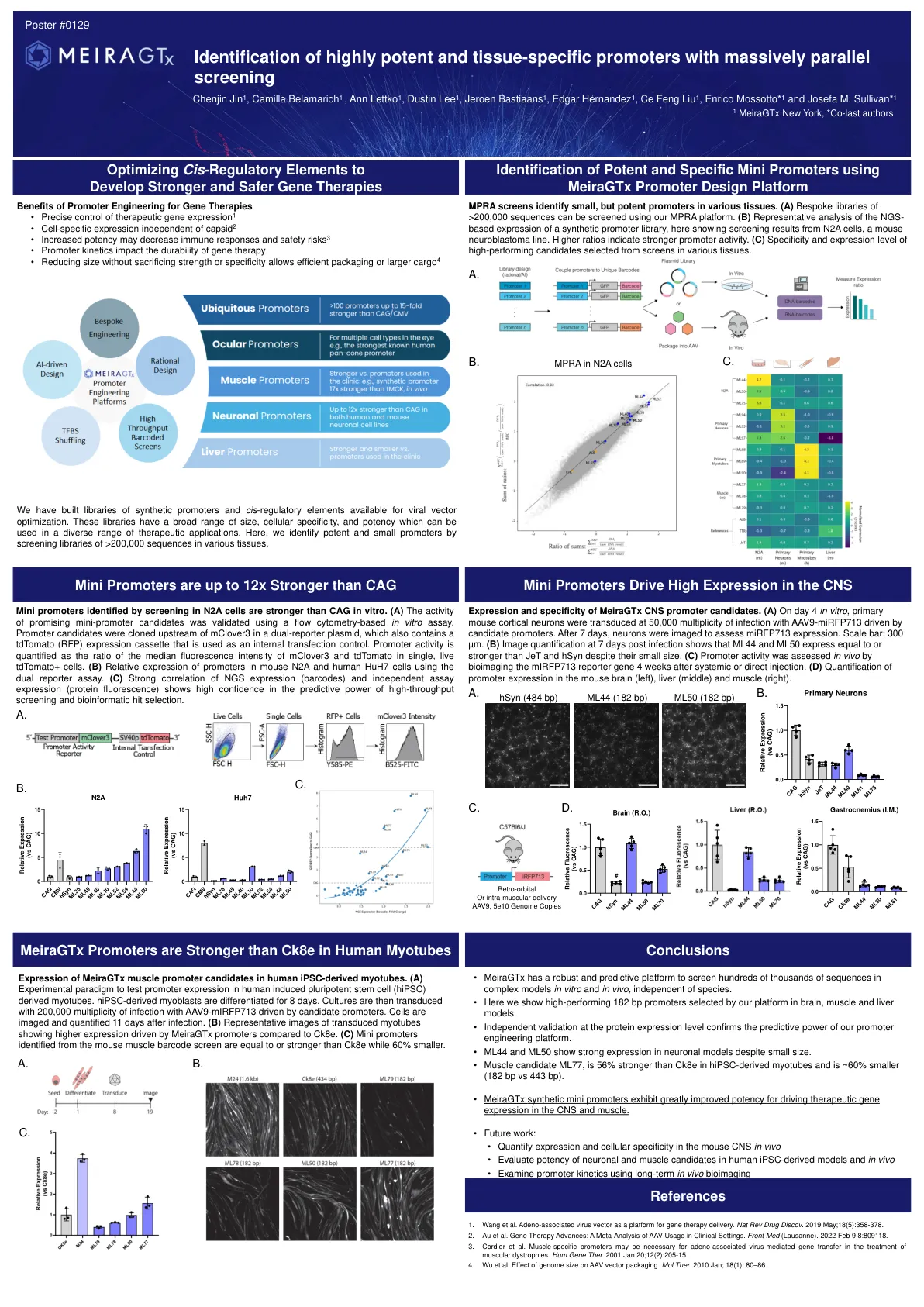
鉴定具有大量平行筛选的高效和组织特异性启动子
迷你启动子在体外比CAG强。(a)使用基于流式细胞术的体外测定法对有希望的迷你启动候选者的活性进行了验证。启动子候选物被克隆在双重孢子质粒中的McLover3上游,该质粒还包含TDTOMATO(RFP)表达盒,该盒被用作内部转染对照。启动子活性被量化为单个活的TDTOMATO+细胞中McLover3和TDTomato的中位荧光强度的比率。(b)使用双报告基因测定法分析,启动子在小鼠N2a和人HuH7细胞中的相对表达。(c)NGS表达(条形码)和独立测定表达(蛋白质荧光)的强相关性表现出对高通量筛选和生物信息学命中选择的预测能力的高信心。

FW:FOI 24/212 mRNA COVID 疫苗、SV40 启动子序列等中存在 DNA 污染 - MHRA 客户服务 - 展望
如果您对内部审查结果不满意,您有权直接向信息专员申请作出决定。请记住,除非您先联系我们进行内部审查,否则信息专员通常不会审查我们对您的请求的处理。您可以通过电子表格在线联系信息专员:https://ico.org.uk/make-a-complaint/foi-and-eir-complaints/foi-and-eir-complaints/

哺乳动物发育增强子激活过程中增强子—启动子相互作用增强
增强子或顺式调控元件可确保在发育过程中对基因表达进行精确的时空控制。该过程由转录因子 (TF) 和辅激活因子介导,它们将调控信息从增强子传递到其目标启动子,跨越的距离可能超过一兆碱基 1-4 。这种增强子-启动子 (E-P) 通讯被认为发生在所谓的拓扑相关结构域 (TAD) 内,拓扑相关结构域是通过黏连蛋白和 CCCTC 结合因子 (CTCF) 的环挤压过程形成的基因组基本组织单位 5-7 。TAD 或 TAD 内染色质相互作用的破坏可能导致基因表达或基因激活的错误下调,并可能导致人类疾病,这表明正确的 E-P 通讯对基因激活的重要性 8-10 。