XiaoMi-AI文件搜索系统
World File Search System噬菌体

某些Vibrio spp的传染性特征。噬菌体...
Vibrio spp。是水产养殖中严重疾病的原因。这项研究旨在开发纤维属。对环境安全友好的抑制方法,即噬菌体(噬菌体)。Vibrio spp。及其噬菌体是从湄公河三角洲省(Kien Giang,Soc Trang和Bac Lieu Provinces)的农场中孤立的。然后,通过比较感染前和感染后的基因序列来研究噬菌体对细菌浓度和TOXR基因的影响。聚合酶链反应鉴定出31株颤音菌株,其中包括9种弧菌偏溶剂。然后使用几个孤立的弧菌属。作为宿主,分离了32个噬菌体菌株。结果发现,每种噬菌体都以特定的方式影响了颤音毒Toxr基因。噬菌体ST9和噬菌体ST10没有改变细菌浓度,但是与副溶血杆菌序列相比,它们可以在4个位置改变核苷酸。然而,噬菌体KG6可以降低细菌浓度,但不会影响基因序列。噬菌体可以通过多种机制影响细菌,包括通过裂解过程降低细菌浓度或影响编码细菌毒力的基因。如果这种作用可以减少或中和细菌的毒力,这些发现在噬菌体治疗研究方面打开了新的方向。
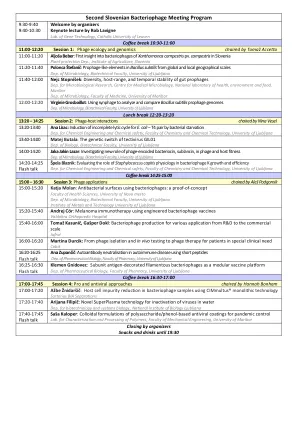
第二斯洛文尼亚噬菌体会议计划
咖啡休息14:25-15:00 15:00 - 16:30 3:噬菌体申请由AlešPodgornik主持,15:00-15:20 Katja Molan:使用噬菌体的抗菌表面:使用新的卫生科学学院证明了Novo MeSto Dep的健康科学。卢布尔雅那大学金属金属学院生物技术学院卢布尔雅那大学生物技术教师15:20-15:40AndrejCör:使用工程性噬菌体疫苗使用工程性噬菌体疫苗的黑色素疗法valdoltra obthopedic Hospital 15:40-16:40-16:00-40-16:00-16:00-tomage for tomage for tomage for tomage for tomage for tomage for tomage for tomage,商业规模的研发Jafral 16:00-16:20 Martina Durcik:从噬菌体隔离和体内测试到特殊临床需求中患者的噬菌体疗法Cobik 16:20-16:25 Flash Talk Talk

噬菌体 - 宿主相互作用作为建立噬菌体在调节发酵食品微生物组成中的作用的平台
抽象食品发酵依赖于健壮的起动培养物的活性,这些培养物通常由乳酸细菌(例如乳酸菌和嗜热链球菌)组成。虽然噬菌体感染代表了可能导致发酵缓慢或失败的持续威胁,但它们在发酵中的有益作用也得到了赞赏。为了发展强大的起动培养物,重要的是要了解噬菌体如何与这些复杂微生物群落的组成景观相互作用并调节。培养依赖性和非依赖性方法对定义许多乳酸细菌(LAB)的单个噬菌体宿主相互作用具有重要作用。需要整合和扩展这些知识,以通过培养物,元基因组学和噬菌体学的结合来充分了解与发酵食品有关的这种相互作用的整体复杂性。有了这样的知识,人们认为可以开发特定于工厂特定的检测和监测系统,以确保强大而可靠的发酵实践。在这篇综述中,我们探索/讨论实验室的噬菌体 - 宿主相互作用,毒和温带噬菌体在微生物组成中的作用以及发酵食品的噬菌体的当前知识。

肌肉骨骼感染中的噬菌体治疗
肌肉骨骼感染(MSI),包括人造关节感染(PJI)和与骨折相关感染(FRI),在现代创伤和骨科手术中仍然是毁灭性的并发症,具有重大的财务和心理成本,并具有严重的财务和心理成本,并增加了发病率(1,2)。PJI包含一系列感染,发生在不同的位置(例如髋关节,膝盖和肩膀)以及不同类型的假体(一线假体或修订假体,包括铰接假体,切除假体,假体 - 关节固定术和股骨全部)(3)。对于PJI患者,有必要避免死前的临床情况,因为经济截肢或髋关节脱落与灾难性功能结果有关。fri可以包括手术和非手术治疗的骨折。对于严重合并症或相关骨缺损的FRI患者中,大约3–17%,截肢是唯一的选择(4)。虽然大多数中心的PJI发生率在0.5%至2%之间(3,5),但周五的发病率从1%到30%,取决于损伤的严重程度(6)。鉴于人口老龄化,脆弱性骨折的数量越来越多,关节置换手术的增加,预计MSI的发生率只会在接下来的几年中增加(3)。与不发展这种并发症的患者相比,这些感染通常会导致功能障碍,活动能力有限和更高的死亡率,从而对患者和医疗保健系统造成严重负担(3,6)。管理概念由联合手术和抗生素治疗方法组成。由于这些感染与生物膜相关,因此治疗通常需要进行多次修订手术。这不仅是昂贵的,而且需要大量的时间和资源,与不开发星期五的患者相比,周五患者的医疗费用高达七倍(7)。

Anti-CRISPR:噬菌体对抗细菌的防御策略
DOI:https://doi.org/10.22271/j.ento.2020.v8.i6n.7968 摘要 成簇的规律间隔的短回文重复序列 (CRISPR)-Cas 系统是一种序列特异性的适应性免疫策略,广泛存在于原核生物系统中,赋予针对各种噬菌体和其他 MGE(如质粒、基因组岛、整合和接合元件)的先天免疫力。即使存在如此复杂的机制,细菌也并非总是能完全战胜噬菌体。这是由于噬菌体和其他 MGE 产生的抗 CRISPR 蛋白。自 2013 年发现以来,迄今为止已鉴定出 60 多个 Acr 家族,还有更多家族尚待发现。研究揭示了 Acrs 采用的多种机制,通过这些机制介导对 CRISPR 防御系统的控制。随着该领域的发展,Acrs 可作为 CRISPR-Cas 技术的潜在控制策略。在本综述中,我们重点介绍了各种抗 CRISPR 蛋白的发现、它们对抗细菌 CRISPR-Cas 系统的作用机制,以及 Acrs 在基因编辑和基因治疗技术中的潜在应用。 关键词:抗 CRISPR、噬菌体、Cas 蛋白、CRISPR、基因组编辑 简介 噬菌体和细菌已经战争了数百万年,噬菌体控制着细菌种群的数量和组成。为了对抗来自噬菌体的这种持续威胁,细菌进化出了非常多样化的防御策略,将检查点设置在噬菌体生命周期的各个阶段。这包括阻断噬菌体附着、抑制 DNA 进入、开发限制-修饰系统、流产感染系统 (Abi) 和干扰噬菌体组装 [1] 。除了上述策略外,细菌还进化出了一种称为 CRISPR Cas 的序列特异性适应性免疫策略 [2] 。 CRISPR 阵列是有关先前感染细菌细胞的噬菌体的数据仓库。Cas 蛋白与 CRISPR 阵列一起构成了这种 RNA 引导的核酸酶复合物。宿主细胞借助称为原间隔区相邻部分 (PAM) 的短序列区分自身 DNA 和入侵的外来移动遗传元件的 DNA [3] 。作为适应性免疫的一部分,所有系统都通过三个主要阶段发挥作用:适应或间隔区获取、表达或生物发生以及干扰阶段。在第一阶段,Cas1-Cas2 复合物识别 PAM 并切除目标 DNA 的一小部分(称为原间隔区),然后将其作为间隔区序列整合到 CRISPR 基因座中。其他辅助因子(如 Cas4、Cas1、Csn2 和逆转录酶 (RT))也可以参与获取阶段。在下一个阶段,CRISPR 基因座转录为单个前 crRNA,然后加工成成熟的 CRISPR RNA (crRNA) [4] 。每个 crRNA 包含部分侧翼重复序列和间隔序列。在干扰阶段,crRNA 与 Cas 蛋白一起被招募以形成核糖核蛋白复合物,该复合物继续在细胞中寻找与 crRNA 间隔序列的任何匹配。如果发现,则根据 CRISPR Cas 系统的类别,通过招募新蛋白质或在复合物本身内激活来启动核酸酶活性。CRISPR-Cas 系统分为两类、六种类型和 30 多个亚型 [5, 6, 7]。第 1 类 CRISPR Cas 系统包括 I、III 和 IV 型,并使用多亚基 Cas 效应分子形成级联复合物。而在第 2 类系统(II、V 和 VI 型)中,靶标识别、结合和切割功能由单个效应蛋白执行。由于这种高度多样化和高效的机制,CRISPR Cas 系统不仅可以保护细菌免受噬菌体的侵害,还可以保护细菌免受其他移动遗传元件 (MGE) 的侵害,如质粒、基因组岛、整合元件和接合元件 [8]。

噬菌体在非洲农业群中的应用
抗菌药物包括用于治疗或预防感染的抗生素,抗病毒药物,抗真菌药和抗寄生虫。对抗菌剂的抵抗是人类的一个问题,但也影响环境,植物和其他动物,包括用于食品生产的动物。一种健康3方法认识到人类,动物和环境的健康之间的相互作用和相互依存关系4。通过考虑使用一个健康镜头的AMR,可以确定新的解决方案,以在全球范围内解决AMR,并解决跨多个领域的实践和过程,从而阻碍了可持续使用抗菌药物。这可能包括改善医疗保健系统和生活水平

解决现代噬菌体疗法的研发差距
抗菌耐药性在全球范围内正在上升,促使噬菌体疗法的研究和发育增加(R&D)作为解决难以处理的细菌感染的一种策略。我们回顾了噬菌体治疗研究的当前状态,包括针对噬菌体研发的主要作战,认知和生物学挑战,并讨论了一些新的方法来开发最近突破性的突破,例如人工智力和合成噬菌体生产。此外,鉴于商业抗菌创新的持续困扰以及当前的公共私人努力,我们将这些研发挑战和挑战背景下,以振兴抗菌药物发现的渠道。我们以反映了在所有收入环境中易于获得新的噬菌体疗法的潜力,以更好地确保患者的访问权限,并考虑到当前公共和公共 - 私人私人解决方案的可能替代方案。

噬菌体 x 右侧操纵子的核苷序列
限制性片段。为了制备微克量的 Hin 375、Hin 550 和 Hae 790(见图 1),将含有示踪量 lambda [32p]_ DNA(2 X 106 cpm)的 5 mg 纯化 lambda DNA 用 Hin(7)或 Hae(6)消化,乙醇沉淀,重悬于 500 ul DNA 缓冲液(5 mM NaCi、10 mM Tris-HCl,pH 7.4、1 mM EDTA)中,在含有 TBE(1)缓冲液的 3.5% 聚丙烯酰胺凝胶(6 mm X 20 cm X 40 cm)上以 320 V 电泳 23 小时。通过放射自显影定位含有适当限制性片段的凝胶部分,切除,并通过苯酚提取去除 DNA(10)。如前所述,从含有 32P 的 DNA 中分离出高比活度标记的限制性片段(2)。通过聚丙烯酰胺凝胶电泳确定每个片段的链长(1、2)。
噬菌体 DNA 分离试剂盒产品说明书
噬菌体 DNA 分离试剂盒产品说明书 产品编号 46800 Norgen 的噬菌体 DNA 分离试剂盒提供了一种快速方法,可从在液体培养的细菌中繁殖的噬菌体中分离和纯化总 DNA。无需使用苯酚、氯仿或氯化铯即可分离 DNA。基于旋转柱的程序速度很快,可在 45 分钟内完成。该试剂盒可高效处理少量噬菌体上清液 (1 mL)。纯化的 DNA 具有最高的完整性,可用于多种下游应用,包括南方印迹、限制性片段长度多态性 (RFLP)、测序、克隆和实时 PCR。Norgen 的纯化技术 纯化基于旋转柱层析。无需使用苯酚、氯仿或氯化铯,即可优先从其他细胞成分(如蛋白质)中纯化噬菌体 DNA。该程序的起始材料是澄清的噬菌体上清液,该上清液已从液体培养物中的细菌碎片中分离出来。首先,使用提供的裂解缓冲液 B 通过热和化学裂解过程裂解噬菌体颗粒(请参阅第 4 页的流程图)。将异丙醇添加到裂解物中,然后将溶液加载到旋转柱上。Norgen 的旋转柱以取决于离子浓度的方式结合核酸,因此只有 DNA 会与柱结合,而大多数 RNA 和蛋白质会在流过中被去除。然后用提供的洗涤溶液 A 洗涤结合的 DNA 以去除任何残留杂质,并用洗脱缓冲液 B 洗脱纯化的总 DNA。纯化的总噬菌体 DNA 具有最高的完整性,可用于许多下游应用。试剂盒组件

利用 CRISPR-Cas13a 进行噬菌体基因组工程
巨型噬菌体(例如铜绿假单胞菌 Ф KZ)具有作为抗菌剂和揭示基本噬菌体生物学模型的潜力。由于蛋白质“噬菌体核”结构可防止 DNA 靶向 CRISPR-Cas 工具的攻击,目前这两种研究都因缺乏基因工程工具而受到限制。为了为 DNA 巨型噬菌体提供反向遗传学工具,我们将同源重组与 RNA 靶向 CRISPR-Cas13a 酶相结合,并使用抗 CRISPR 基因 (acrVIA1) 作为可选择标记。我们表明,该过程可以插入外来基因、删除基因并向 Ф KZ 基因组添加荧光标签。内源性 gp93 的荧光标记显示它会随噬菌体 DNA 一起排出,而微管蛋白样蛋白 PhuZ 的缺失令人惊讶地对噬菌体爆发大小的影响很小。还成功编辑了另外两种能够抵抗 DNA 靶向 CRISPR-Cas 系统的噬菌体。RNA 靶向 Cas13a 有望成为难治性噬菌体的通用基因编辑工具,从而能够系统地研究功能未知的噬菌体基因。