XiaoMi-AI文件搜索系统
World File Search System增强子
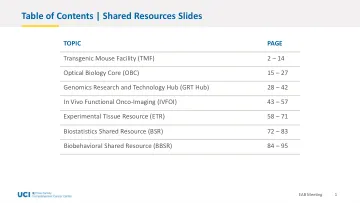
目录 | 共享资源幻灯片
• 由于缺乏有效的体内模型,与人类遗传病相关的非编码变异的功能表征仍然具有挑战性。 • Dual-enSERT-2 是一种强大的基于 CRISPR 的双色荧光报告系统,可快速定量分析活体 G0 第一代转基因小鼠中的增强子等位基因活性。 • Dual-enSERT 可与单细胞转录组学相结合,以细胞分辨率表征变异增强子等位基因活性,揭示与致病增强子失调有关的候选分子通路(例如介导神经胶质瘤形成的 IDH1 增强子变异)。 • TMF 对小鼠受精卵和胚胎进行显微注射和电穿孔,并将其转移到假孕受体小鼠中供 Kvon 实验室使用。

远距离连接:绘制神经细胞染色质中全基因组长距离相互作用图谱,以识别神经发育障碍的新候选基因
摘要:与神经发育障碍 (NDD) 和特征相关的 DNA 序列变异(单核苷酸多态性或变异,SNP/SNV;拷贝数变异,CNV)通常映射到假定的转录调控元件上,特别是增强子。然而,这些增强子控制的基因仍然定义不清。传统上,给定增强子的活性及其与序列变异相关的可能改变的影响被认为会影响最近的基因启动子。然而,在神经细胞染色质中获得全基因组长距离相互作用图挑战了这种观点,表明给定的增强子通常不与最近的启动子相连,而是与更远的启动子相连,跳过中间的基因。在本篇观点中,我们回顾了一些最近的论文,这些论文生成了长距离相互作用图谱(通过 HiC、RNApolII ChIA-PET、Capture-HiC 或 PLACseq),并将已识别的长距离相互作用 DNA 片段与与 NDD(如精神分裂症、躁郁症和自闭症)和特征(智力)相关的 DNA 序列变体重叠。这种策略允许将承载 NDD 相关序列变体的增强子的功能归因于位于线性染色体图谱远处的连接基因启动子。其中一些增强子连接基因确实已被鉴定为导致疾病,通过鉴定基因蛋白质编码区(外显子)内的突变,验证了该方法。然而,重要的是,连接基因还包括许多以前未在其外显子中发现突变的基因,指向 NDD 和特征的新候选贡献者。因此,长距离相互作用图谱与检测到的与 NDD 相关的 DNA 变异相结合,可用作识别新的候选疾病相关基因的“指针”。基于 CRISPR-Cas9 的方法对涉及增强子和启动子的长距离相互作用网络进行功能操控,开始探索已识别相互作用的功能意义以及所涉及的增强子和基因,从而提高我们对神经发育及其病理学的理解。

一个共享的古老增强子元件差异地调节果蝇腿部发育过程中的 bric-a-brac 串联基因重复
基因复制和转录增强子的出现/修饰被认为对动物进化过程中表型创新做出了巨大贡献。尽管如此,人们对基因复制后增强子如何进化以及调控信息如何在复制基因之间重新连接知之甚少。果蝇 bric-a-brac (bab) 复合体由串联旁系同源基因 bab1 和 bab2 组成,为解决这些问题提供了范例。我们之前描述了一种调节发育足中 bab2 表达的基因间增强子 (名为 LAE)。我们在此显示直接与 LAE 结合的 bab2 调节子也控制跗骨细胞中的 bab1 表达。通过 CRISPR/Cas9 介导的基因组编辑切除 LAE 表明,这种增强子似乎参与了 bab1 和 bab2 在腿部组织中共表达,但并不是严格必需的。相反,LAE 增强子对于沿近端-远端足轴的旁系同源物特异性 bab2 表达至关重要。染色质特征和表型挽救实验表明,LAE 功能部分冗余,腿特异性调控信息与 bab1 转录单元重叠。系统基因组学分析表明 (i) bab 复合体起源于 Cyclorrhapha dipteran 亚系早期祖先单基因的复制,以及 (ii) LAE 序列在 Brachycera 亚目中很早就已进化固定,因此早于基因复制事件。这项工作为增强子提供了新的见解,特别是关于它们的出现、维持和进化过程中的功能多样化。

鉴定在哺乳动物中具有活性的祖先有颌类 Gli3 增强子
已知转录调节因子和 Hh 信号通路效应因子 Gli3 的异常表达会引发先天性疾病,最常影响中枢神经系统 (CNS) 和四肢。准确描绘胚胎发育过程中控制 Gli3 转录的基因组顺式调控景观对于解释与先天性缺陷相关的非编码变异至关重要。在这里,我们对分子进化速度较慢的鱼类进行了比较基因组分析,以识别 Gli3 内含子间隔 (CNE15-21) 中七个以前未知的保守非编码元件 (CNE)。斑马鱼的转基因试验表明,这些元件中的大多数驱动 Gli3 表达组织中的活动,主要是鳍、中枢神经系统和心脏。这些 CNE 与人类疾病相关的 SNP 的交集确定了 CNE15 是一种假定的哺乳动物颅面增强子,在脊椎动物中具有保守活性,并且可能受到与人类相关的突变的影响

鉴定免疫相关增强子RNA SATB1-AS1是胸腺癌预后的新型生物标志物
。CC-BY 4.0国际许可证可永久提供。是作者/资助者,他已授予Medrxiv的许可证,以显示预印本(未通过PEER REVIVE的认证)Preprint Preprint the版权所有此版本,该版本于2025年2月14日发布。 https://doi.org/10.1101/2025.02.13.25322204 doi:medrxiv preprint

评估M/ ... div>的长短期限内存网络
抽象增强器协调驱动多细胞发展和谱系承诺的基因表达程序。因此,人们认为增强子的遗传变异通过改变细胞命运承诺会导致发育疾病。然而,尽管已经确定了许多含有变异的增强子,但缺乏内生测试这些增强剂对谱系承诺的影响的研究。我们执行一个单细胞CRISPRI筛选,以评估与先天性心脏缺陷(CHD)有关的25种增强子和推定心脏靶基因的内源性作用。我们确定了16个增强剂,其抑制导致人类心肌细胞(CMS)的分化不足。专注的CRISPRI验证屏幕表明,TBX5增强剂的抑制延迟了从中期到后期CM状态的转录开关。两个TBX5增强剂表观遗传扰动的内源性遗传缺失。共同确定了心脏发育的关键增强子,并表明这些增强剂的不正调可能导致人类患者的心脏缺陷。

通过靶向增强子启动途径在患者来源的胰腺癌细胞中进行启动治疗
a 马赛癌症研究中心 (CRCM)、INSERM U1068、CNRS UMR 7258、Luminy 科学与技术公园、艾克斯-马赛大学和保利-卡尔梅特研究所,法国 b 布宜诺斯艾利斯大学、国家科学技术研究委员会、药理学和植物学研究中心 (CEFYBO)、医学院,布宜诺斯艾利斯,阿根廷 c 布宜诺斯艾利斯大学、医学院、微生物学、寄生虫学和免疫学系,布宜诺斯艾利斯,阿根廷 d 肿瘤身份证计划 (CIT)、法国抗癌联盟,巴黎,法国 e Laboratoire Modal ' X - UMR 9023,巴黎南泰尔大学,法国南泰尔 f 巴黎萨克雷大学、AgroParisTech、INRAE、UMR MIA Paris-Saclay, Palaiseau 91120,法国 g 基因组学和精准医学中心(GSPMC),威斯康星医学院,美国威斯康星州密尔沃基 h 威斯康星医学院外科系研究部,美国威斯康星州密尔沃基 i 雷恩大学,CNRS,INSERM,IGDR(雷恩遗传和发展研究所)- UMR 6290,ERL U1305,雷恩,法国 j 巴黎城大学,炎症研究中心(CRI),INSERM,U1149,CNRS,ERL 8252,巴黎 F-75018,法国 k 埃尔克鲁塞阿尔塔综合医院,Florencio Varela,文学士,阿根廷 l 阿图罗·豪雷切大学,Florencio Varela,文学士,阿根廷

一套增强子 AAV 和转基因小鼠系,用于 1 基因访问皮质细胞类型 2
Yoav Ben-Simon, 1,4 Marcus Hooper, 1,4 Sujatha Narayan, 1,4 Tanya Daigle, 1,4 Deepanjali Dwivedi, 1 Sharon W. 4 Way, 1 Aaron Oster, 1 David A. Stafford, 2 John K. Mich, 1 Michael J. Taormina, 1 A. Refugio, 1 A. Martina-Jamena. R. Roth, 1 Shona Allen, 2 Angela Ayala, 1 Trygve E. Bakken, 1 Tyler Barcelli, 1 Stuard Barta, 1 6 Jacqueline Bendrick, 1 Darren Bertagnolli, 1 Jessica Bowlus, 1 Gabriella Boyer, 1 Krissy Brouner, 1 Brittny Casian, 1 7 Chara Chair, Chara Rush, 1 Chara Rush. barty, 1 Rebecca K. Chance, 2 Sakshi Chavan, 1 Maxwell 8 Departee, 1 Nicholas Donadio, 1 Nadezhda Dotson, 1 Tom Egdorf, 1 Mariano Gabitto, 1 Jazmin Garcia, 1 Amanda 9 Gary, 1 Molly Gasperini, 1 Jeffry Goldy, 1 1 Blanche, 1 Lucas Gregory, No. . 1 Francoise Haeseleer, 1 10 Carliana Halterman, 1 Olivia Helback, 1 Dirk Hockemeyer, 2 Cindy Huang, 1 Sydney Huff, 1 Avery Hunker, 1 Nelson 11 Johansen, 1 Zoe Juneau, 1 Brian Kalmbach, 1 Shannon Khem, 1 Emily Kuckel, 1 Lar Rasen, 1 12 Changkyu Lee, 1 Angus Y. Lee, 2 Madison Leibly, 1 Garreck H. Lenz, 1 Elizabeth Liang, 1 Nicholas Lusk, 1 Jocelin 13 Malone, 1 Tyler Mollenkopf, 1 Elyse Morin, 1 Dakota Newman, 1 Lydia Ng, 1 Kiet Ngoste, 1 1 Victoria Oman, 14 h Pham, 1 Christina A. Pom, 1 Lydia Potekhina, 1 Shea Ransford, 1 Dean Rette, 1 Christine 15 Rimorin, 1 Dana Rocha, 1 Augustin Ruiz, 1 Raymond EA Sanchez, 1 Adriana Sedeno-Cortes, 1 Joshua P. Sevigny, 1 Nadi Lava, 16 Lyvalomi Ana R. Sigler, 1 La' Akea Siverts, 1 Saroja Somasundaram, 1 Kaiya 17 Stewart, 1 Eric Szelenyi, 1 Michael Tieu, 1 Cameron Trader, 1 Cindy TJ van Velthoven, 1 Miranda Walker, 1 Natalie 18 Weed, 1 Morgan Wirlin, 1 Toren Wood, 1 Toren Wood, 1 Zilda o, 1 Thomas Zhou, 1 Jeanelle Ariza, 1 Nick 19 Dee, 1 Melissa Reding, 1 Kara Ronellenfitch, 1 Shoaib Mufti, 1 Susan M. Sunkin, 1 Kimberly A. Smith, 1 Luke 20 Esposito, 1 Jack Waters, 1 Bargavi Thyagarajan, 1 Yaqin , 1 Shenq , 1 Sheng Leng . Boaz P. Levi, 1 John 21 Listen, 2,3 Jonathan Ting, 1 Bosiljka Tasic 1,5,* 22

利用深度学习模型基于基因组和表观基因组特征来识别增强子和超级增强剂
抽象的超级增强剂是转录因子和染色质调节剂密集地占据的活性超增强剂的类别,可控制疾病相关基因的表达和细胞身份。最近的研究表明,各种因素和超级增强剂,尤其是在各种癌症中形成复杂结构。然而,我们目前对超级增强剂的了解,例如其基因组位置,与其他超级增强剂区域的因素,功能和区别的相互作用仍然有限。这项研究旨在采用深度学习技术来基于基因组和表观基因组特征来检测和区分超级增强剂和增强剂,并将结果的准确性与本研究中的其他机器学习方法进行比较,除了评估算法外,我们还培训了一组基因组和表观基因组和表观学算法 - DNA序列中的超级增强剂。我们以更高的精度和精度成功地预测了序列中超级增强剂的存在。

双向表观遗传编辑揭示基因调控的层次结构
CRISPR 扰动是研究基因组功能效应的宝贵工具。然而,现有方法在研究非编码元件和遗传相互作用方面的效用有限。在这里,我们开发了一个双向表观遗传编辑系统 (CRISPRai),其中正交激活 (CRISPRa) 和抑制 (CRISPRi) 扰动同时应用于同一细胞的多个基因座。我们开发了双 gRNA 捕获单细胞 Perturb-seq 来研究两种造血谱系转录因子 SPI1 和 GATA1 之间已建立的相互作用,并发现了共同调节基因的新型上下文特定调控模式。将 CRISPRai 扩展到非编码元件,我们解决了多个增强子如何相互作用以调节 T 细胞中共同靶基因白细胞介素-2 的表达。我们发现增强子功能主要是附加的并能够对基因表达进行微调,但在基因表达控制强度方面,增强子之间存在明显的层次结构。启动子在控制基因表达方面比大多数增强子占主导地位;然而,一小部分增强子表现出强大的功能效应或守门人功能,尽管启动子被激活,但仍可以关闭基因。将这些功能数据与组蛋白 ChIP-seq 和 TF 基序富集相结合,表明存在多种增强子介导的基因调控模式。我们的方法 CRISPRai 用于双向表观遗传编辑,提供了一种识别新遗传相互作用的方法,这些相互作用在没有双向扰动的情况下进行研究时可能会被忽视,并且可以应用于基因和非编码元件。