XiaoMi-AI文件搜索系统
World File Search System核酸酶
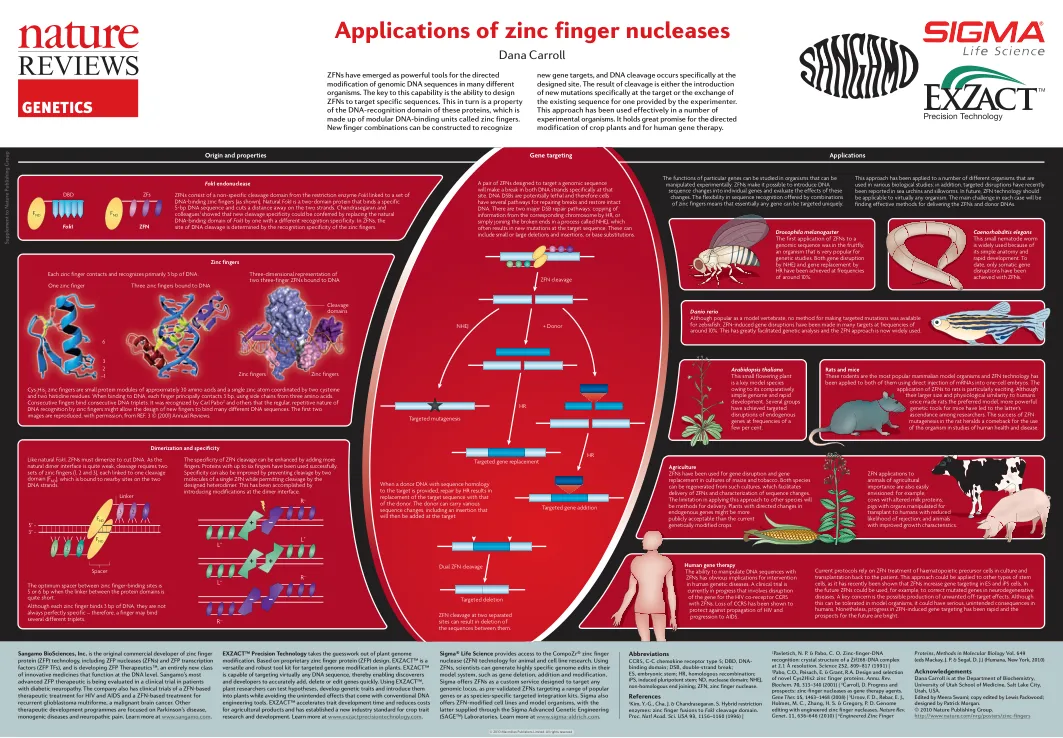
锌指核酸酶的应用
EXZACT™ 精准技术消除了植物基因组改造中的猜测。EXZACT™ 基于专有的锌指蛋白 (ZFP) 设计,是一种多功能且强大的工具包,可用于对植物进行靶向基因组改造。EXZACT™ 能够针对几乎任何 DNA 序列,从而使发现者和开发者能够快速准确地添加、删除或编辑基因。使用 EXZACT™,植物研究人员可以测试假设、开发遗传特性并将其引入植物,同时避免传统 DNA 工程工具带来的意外影响。EXZACT™ 加快了特性开发时间并降低了农产品成本,并为作物特性研发建立了新的行业标准。了解更多信息,请访问 www.exzactprecisiontechnology.com。
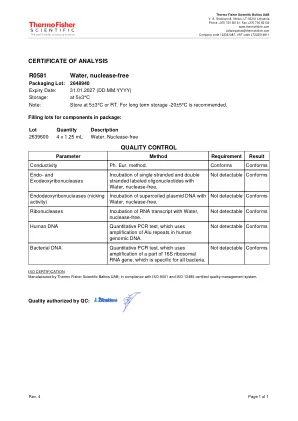
R0581水,无核酸酶
R0581水,无核酸酶包装批:2648940到期日期:31.01.2027(dd.mm.yyyy)存储:在5±3°C下注:存储在5±3°C或RT。建议长期存储-20±5°C。

ArciTect™ Cas9-eGFP 核酸酶
ArciTect™ Cas9-eGFP 核酸酶是一种融合蛋白,由增强型绿色荧光蛋白 (eGFP) 和来自化脓性链球菌的野生型 Cas9 重组蛋白组成;它包含一个 C 端连接的 eGFP 分子。ArciTect™ Cas9-eGFP 核酸酶需要与向导 RNA(例如 ArciTect™ sgRNA(目录号 #200-0013)或由 ArciTect™ tracrRNA(目录号 #76016)和 ArciTect™ crRNA(目录号 #76010)组成的双链)结合,以形成核糖核蛋白 (RNP) 复合物。该 RNP 复合物在基因组中的位点特定位置产生双链断裂。 ArciTect™ Cas9-eGFP 核酸酶还在 N 端含有核定位信号,确保 RNP 复合物转位至细胞核,从而提高基因组编辑的效率。由于 RNP 复合物在转染后完全发挥作用,因此在转位至细胞核后可立即发挥作用。RNP 复合物在 48 小时内降解,为基因组编辑提供了充足的时间,同时减少了 RNP 复合物持续存在可能导致的脱靶效应。使用 RNP 系统还可以避免生成稳定的 Cas9 表达细胞系的繁琐过程,从而节省时间并降低由于可诱导表达系统泄漏而导致脱靶效应的风险。化脓性链球菌 Cas9 使用原间隔区相邻基序 (PAM) 序列 NGG(其中 N 可以是任何核苷酸)。如果靶序列下游没有基因组 PAM 位点,酶就不会裂解。

Lb Cas12a 核酸酶
免责声明:在法律允许的范围内,赛默飞世尔科技公司和/或其附属公司不对与本文件有关或由本文件引起的(包括您对其的使用)的特殊、偶然、间接、惩罚性、多重或后果性损害承担责任。

特定的弧菌核酸酶,以消除
本演示文稿(以及我们在此处可能与我们有关的任何其他陈述或信息)包含1995年《私人证券诉讼改革法》的含义中的前瞻性陈述。All statements contained in this presentation (together with any other statements or information that we may make in connection herewith) that do not relate to matters of historical fact should be considered forward-looking statements, including, without limitation, statements regarding the development of our product candidates involving our ARCUS® genome editing platform, our ARCUS-HBV nuclease and our in vivo gene editing product candidates including PBGENE-HBV and expected 2024年的里程碑。在某些情况下,您可以通过诸如“目标”,“预期”,“实现”,“相信”,“思考”,“可以”,“估算”,“期望”,“期望”,“目标”,“预期”,“五月”,“五月”,“五月”,“使命”,“任务”,“计划”,“预测”,“预测”,“优势”,“ project”,“”,“”,“”,“”或否定的词和类似的词和表达。前瞻性陈述基于管理层当前的期望,信念和假设以及当前可用的信息。

nzy核糖核酸酶抑制剂
nzy核糖核酸酶抑制剂是一种从大肠杆菌中纯化的重组蛋白。它通过以1:1的比例非共归因于胰腺类型(例如RNase A,RNase B和RNase C)抑制胰腺类型的核糖核酸(RNase; EC 3.1)的活性。nzy核糖核酸酶抑制剂在RNase污染是潜在问题的任何应用中都是有用的。例如,它可用于保护cDNA合成反应,RT-PCR或体外转录/翻译中的模板RNA,并在体外复制过程中保护病毒RNA。此外,它将在RNA分离和纯化和无RNase抗体制备过程中抑制RNase。nzy核糖核酸酶抑制剂对RNase 1,RNase T1,RNase T2,S1核酸酶和RNase H.

通过微型DNA核酸酶ISCB
预印本(未通过同行评审认证)是作者/资助者。保留所有权利。未经许可就不允许重复使用。此版本的版权持有人于2025年2月14日发布。 https://doi.org/10.1101/2025.02.11.637773 doi:Biorxiv Preprint

核酸酶和切口酶基因修饰的改进......
基因组编辑技术的进步使得利用酶的功能进行有效的 DNA 修饰成为可能,这对治疗人类遗传疾病具有巨大的潜力。已经开发出几种核酸酶基因组编辑策略来纠正基因突变,包括大核酸酶 (MN)、锌指核酸酶 (ZFN)、转录激活因子样效应核酸酶 (TALEN) 和成簇的规律间隔短回文重复序列-CRISPR 相关蛋白 (CRISPR-Cas)。CRISPR-Cas 已被进一步设计为创建切口酶基因组编辑工具,包括具有高精度和高效率的碱基编辑器和主要编辑器。在这篇综述中,我们总结了用于治疗遗传疾病的核酸酶和切口酶基因组编辑方法的最新进展。我们还强调了这些方法转化为临床应用的一些局限性。

nuc-off核酸酶和DNA去除喷雾
图1。在去除RNase和dNase中,MP生物医学Nuc-Off核酸酶和DNA去除喷雾剂和竞争者T溶液的性能比较。A. RNase消除。在室温下孵育5分钟,将4μl的去除试剂和不同量的RNase(以1μl为单位)的混合物孵育;之后,加入1μlRNA,并在室温下进一步孵育15分钟,然后在含有甲醛的琼脂糖凝胶中变性和最终混合物的电泳。B. DNase消除。在室温下孵育4μl的去除试剂和不同量的DNase(以1μl)的混合物5分钟;之后,将1μl10X反应缓冲液和1μgDNA和无核酸酶的水加入总体积10μl,并在室温下进一步孵育15分钟,然后是最终混合物的琼脂糖凝胶电泳。C.去除试剂对DNA稳定性的影响。在室温下孵育15分钟,将4μl的去除试剂和1μl基因组DNA的混合物进行孵育,然后通过琼脂糖凝胶电泳进行分析。D.去除试剂对RNA稳定性的影响。在室温下孵育4μl的去除试剂和1μlRNA的混合物,然后变性添加含有甲醛的琼脂糖凝胶电泳。此处显示的图仅供参考,它可能会根据不同的实验条件而有所不同。

Cas9 核酸酶 GFP NLS 蛋白
产品描述 成簇的规则间隔短回文重复序列 (CRISPR)/Cas9 系统是基因组编辑中最新的 RNA 引导核酸内切酶工具,可实现非常具体的基因组破坏和替换。Cas9 核酸酶 NLS 与 GFP 的融合可实现转染的视觉确认以及随后的 Cas9 从细胞中清除的验证。Cas9 核酸酶-GFP 也可用于 FACS 应用和筛选。Cas9 核酸酶-GFP NLS 在蛋白质的 C 端包含 SV40 T 抗原核定位序列 (NLS)。