XiaoMi-AI文件搜索系统
World File Search System熔化温度

链球菌突变体调节细菌 - 真菌相互作用中白色念珠菌的泛素修饰
植物病毒对全球农业构成了重大威胁,并需要有效的工具才能及时检测。我们提出了AutoPvprimer,这是一种创新的管道,该管道整合人工智能(AI)和机器学习以加速植物病毒引物的发展。管道使用Biopython从NCBI数据库自动检索不同的基因组序列,以增加后续引物设计的鲁棒性。design_-primers_with_tuning模块使用随机森林分类器,可优化参数并为不同的实验条件提供灵活性。质量控制措施,包括评估Poly-X含量和熔化温度,提高了引物的可靠性。AUTOPVPRIMER独有的是Visualize_primer_dimer模块,它支持引物二聚体的可视化评估,这是其他工具中缺少的功能。引物特异性通过引物爆炸验证,这有助于管道的整体效率。AutoPvprimer已成功地应用于番茄镶嵌病毒,证明其适应性和效率。模块化设计允许用户自定义,并将适用性扩展到不同的植物病毒和实验场景。管道代表了引物设计的重大进展,并为研究人员提供了加速分子生物学实验的有效工具。未来的发展旨在扩展兼容性并纳入用户反馈,以巩固AutoPvprimer,作为对生物信息学工具箱的创新贡献,也是提高植物病毒学研究的有希望的资源。

alcl3-nacl-Zncl2二级电解质在下一步 -
摘要:增加了从例如光伏和风能中存储间歇性可再生电力的需求,导致大量的大规模固定能量存储中的大量研发,例如,斑马电池(Na-Nicl 2固体电解质电池)。用丰富和低成本的Zn代替Ni,使斑马电池更具成本效益。然而,很少对此下一代斑马(Na-Zncl 2)电池系统进行研究,尤其是在其ALCL 3 -NACL-ZNCL 2二级电解质上。其特性(例如相图和蒸气压力)对于细胞设计和优化至关重要。在我们以前的工作中,一种用于熔融盐电解质选择的模拟辅助方法显示了其在熔融盐电池开发中的成功应用。此处使用相同的方法来研究ALCL 3 -NACL-ZNCL 2盐电解质的相图和通过事实TM和热分析技术(差速器扫描量热法(DSC)和最佳电池效果及其对电池性能的影响和放电机制的影响,其相位图和蒸气压力(差分扫描量热法(DSC)和效果。DSC和Optimelt结果表明,诸如熔化温度和相变的实验数据与模拟相图非常吻合。此外,事实TM模拟表明,随着ALCL 3的温度和摩尔分数的升高,盐蒸气压力显着增加。获得的相图和蒸气压将用于辅助电解质选择,电池设计和电池操作。
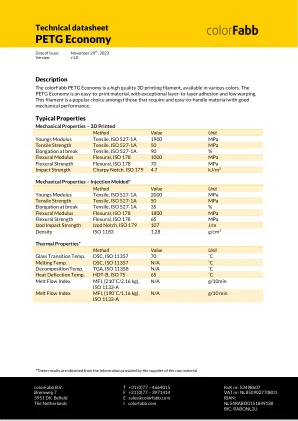
技术数据表PETG经济
机械性能 - 3D印刷方法值单位年轻人模抗拉伸,ISO 527-1A 1900 MPA拉伸强度拉伸,ISO 527-1A 50 MPA在休息拉伸时伸长时527-1A,ISO 527-1A 90%弯曲屈曲屈曲,ISO 1000 MPA 1000 MPA FELTURAL ISO FORTURAL ESO 179 70 70 70 70 M. 4.7 kJ/m 2 Mechanical Properties – Injection Molded* Method Value Unit Youngs Modulus Tensile, ISO 527-1A 2000 MPa Tensile Strength Tensile, ISO 527-1A 50 MPa Elongation at break Tensile, ISO 527-1A 35 % Flexural Modulus Flexural, ISO 178 1800 MPa Flexural Strength Flexural, ISO 178 65 MPa Izod Impact Strength Izod Notch,ISO 179 107 J/M密度ISO 1183 1.28 g/cm 3热性能*方法值单位玻璃透过温度。DSC,ISO 11357 70°C熔化温度。DSC,ISO 11357 N/A°C分解温度。TGA,ISO 11358 N/A°C热偏转温度。HDT-B,ISO 75 65°C熔体流量指数MFI(210˚C/2.16 kg),ISO 1133-A

减少 pegRNA 内固有的自抑制相互作用可提高主要编辑效率
Prime 编辑系统能够在不引入双链断裂的情况下在基因组内进行精确编辑。先前的研究根据序列组成定义了 pegRNA 的最佳引物结合位点 (PBS) 长度约为 13 个核苷酸。然而,最佳 PBS 长度表征是基于使用质粒或慢病毒表达系统的 Prime 编辑结果。在本研究中,我们证明,对于 Prime 编辑器 (PE) 核糖核蛋白复合物,PBS 和间隔序列之间的自抑制相互作用会影响 pegRNA 结合效率和靶标识别。通过降低 PBS-间隔区之间的互补性来破坏这种自抑制相互作用可提高多种 Prime 编辑格式中的 Prime 编辑效率。对于末端保护的 pegRNA,在哺乳动物细胞中,较短的 PBS 长度且 PBS-靶标链熔化温度接近 37 ◦ C 是最佳的。此外,在 PE-pegRNA 递送后对细胞进行短暂冷休克处理可进一步提高具有优化 PBS 长度的 pegRNA 的 prime 编辑结果。最后,我们表明使用这些改进的参数设计的 pegR-NA 编程的 prime 编辑器核糖核酸蛋白复合物可有效纠正患者来源的成纤维细胞中的疾病相关基因突变,并有效地在原代人类 T 细胞和斑马鱼中安装精确编辑。
![arxiv:2502.09044v1 [nucl-th] 2025年2月13日](/simg/7\753cc4731c90c6aea1248f305961a4a2ab993375.webp)
arxiv:2502.09044v1 [nucl-th] 2025年2月13日
结合状态的性质是辐射光谱的基础,并且在从腺体物质到夸克 - 杜松等离子体(QGP)的过渡中起着核心作用。在强耦合QGP(SQGP)中,温度,结合能和较大的碰撞宽度的相互作用在评估HADRONIC状态及其最终熔化的中等内部性能方面带来了巨大的挑战。尤其是,QGP中繁重的Quarkonia的存在是一个长期的问题,很难通过考虑其在真能轴上的光谱特性来解决。我们通过分析复杂能量平面中的中等热力学夸克t- t-含量来解决这个问题。我们首先在真空中验证这种方法,其中很容易识别观察到的状态的t -matrix极。将这种方法部署到QGP中最近计算出的T型t-Matrices中时,我们发现复杂能平面中的极点可以持续到令人惊讶的较大温度,这取决于中等相互作用的强度。虽然精确地定义了极点位置的质量和宽度,但结合能的概念并不是由于缺乏由基础抗/Quark光谱函数的(大)宽度引起的阈值。因此,我们的方法提供了一种新的严格量子力学标准,以确定SQGP中强烈态温度的熔化温度,同时提高了传输参数的理论确定的准确性。

基于脂质纳米粒子的核糖核蛋白递送用于体内基因组编辑
成簇的规则间隔短回文重复序列 (CRISPR) 相关 (Cas) 系统是一种通过 DNA 修复机制进行位点特异性基因破坏、修复和基因组 DNA 修饰的技术,有望成为治疗传染病和遗传疾病的基本治疗策略。对于临床应用,基于非病毒载体的 CRISPR/Cas 核糖核蛋白 (RNP) 递送非常重要,但递送效率低和缺乏实用的制造方法仍然是一个问题。我们在此报告了一种基于脂质纳米颗粒 (LNP) 的 Cas RNP 递送系统的开发,该系统基于优化设计的单链寡核苷酸 (ssODN),可实现高效的体内基因组编辑。序列特异性 RNP-ssODN 复合物的形成被发现对于 RNP 的功能性递送很重要。此外,sgRNA 和 ssODN 之间的熔化温度 (Tm) 对体内基因敲除效率有显著影响。具有高 Tm 的 ssODN 导致有限的敲除 (KO) 活性,而接近室温的 ssODN 显示出最高的 KO 活性,这表明 RNPs 的细胞质释放非常重要。连续两次静脉注射 Tm 优化的配方分别在 DNA 和蛋白质水平上实现了约 70% 和 80% 的转甲状腺素蛋白 KO,且没有任何明显的毒性。这些发现对安全的体内 CRISPR/Cas RNP 递送技术的开发及其在基因组编辑疗法中的实际应用具有重要贡献。

管壳式潜热储热换热器的快速简化模型及其在非线性规划成本优化设计中的应用
1-D PCM 棒的横截面积,[m 2 ] 比热,[J kgK ⁄ ] 运行成本,[$ yr ⁄ ] 电价,[$ kWhr ⁄ ] 管材成本,[$ kg ⁄ ] PCM 材料成本,[$ kg ⁄ ] 管内传热系数,[W m 2 K ⁄ ] 总时间步数 电导率,[W mK ⁄ ] 管总长度,[m ] 平准化能源成本,[$ MWh ⁄ ] PCM 潜能,[kJ kg ⁄ ] 径向网格数 管长网格数 努塞尔特数 普朗特数 传热速率,[W] 传热速率,[W] HTF 总质量流速,[kg s ⁄ ] 环内半径,[m] 环状几何中的移动凝固前沿,[m]环形圆柱体 PCM 的热阻,[ m ] 圆柱体 PCM 内的热阻,[ KW ⁄ ] 导热流体内的热阻,[ KW ⁄ ] 雷诺数 温度,[ ℃ ] 边界冷却温度,[ ℃ ] 相变材料熔化温度,[ ℃ ] 管与圆柱体 PCM 之间的界面温度,[ ℃ ] 管内导热流体的速度,[ ms ⁄ ] 管壁厚度,[ mm ] 壳体厚度,[ mm ] 一维 PCM 棒的长度,[ m ] 每天运行小时数,[ hr ] 凝固时间,[ hr ] 移动凝固前沿,[ m ] 设备总寿命,[ yr ] 环形圆柱体 PCM 的轴长,[ m ] 两个坐标系之间的凝固前沿比率 密度,[ kg m 3 ⁄ ] 粘度,[ Pa ∙s ] 潜能储存系统的有效性矩形几何结构显热能分数因子 圆柱形几何结构显热能分数因子 差值或增量步长 泵效率
mytaqtm DNA聚合酶
重要的考虑因素和PCR优化,最佳条件将从反应到反应,并取决于所使用的模板/引物。5x mytaq反应缓冲液:5x MyTAQ反应缓冲液包括5mm DNTP,15mm MGCL 2,稳定器和增强剂。每个组件的浓度已得到广泛优化,从而减少了进一步优化的需求。其他PCR增强剂,例如HISPEC,Polymate或Betaine等。。引物:正向和反向引物通常以0.2-0.6M的最终浓度使用。我们建议使用0.4M作为最终浓度(即每50 le反应体积的每个引物的下午20点)。 过高的底漆浓度可以降低启动的特异性,从而导致非特异性产物。 设计底漆时,我们建议使用Primer-Design软件,例如Primer3(http://frodo.wi.mit.edu/primer3)或可单位的10mm和3mm和3mm的阳离子阳离子浓度,或者单位阳离子浓度和分别为10mm和3mm。 引物应具有约60°C模板的熔化温度(TM):反应中的模板量主要取决于所使用的DNA类型。 对于低结构复杂性(例如质粒DNA)的模板,我们建议使用50pg-10ng DNA每50°L反应体积。 对于真核基因组DNA,我们建议每50 l反应的起始量为200ng DNA,这可以在5ng-5ng-500ng之间变化。 避免在含EDTA的解决方案中重新悬浮模板(例如)很重要 te buffer)由于EDTA螯合免费Mg 2+。每50 le反应体积的每个引物的下午20点)。过高的底漆浓度可以降低启动的特异性,从而导致非特异性产物。设计底漆时,我们建议使用Primer-Design软件,例如Primer3(http://frodo.wi.mit.edu/primer3)或可单位的10mm和3mm和3mm的阳离子阳离子浓度,或者单位阳离子浓度和分别为10mm和3mm。引物应具有约60°C模板的熔化温度(TM):反应中的模板量主要取决于所使用的DNA类型。对于低结构复杂性(例如质粒DNA)的模板,我们建议使用50pg-10ng DNA每50°L反应体积。对于真核基因组DNA,我们建议每50 l反应的起始量为200ng DNA,这可以在5ng-5ng-500ng之间变化。避免在含EDTA的解决方案中重新悬浮模板(例如te buffer)由于EDTA螯合免费Mg 2+。初始变性:对于非复合模板,例如质粒DNA或cDNA,建议在95°C下1分钟的初始变性步骤。对于更复杂的模板,例如真核基因组DNA,为了促进DNA的完全融化,需要更长的初始变性时间至3分钟。变性:我们的协议建议在95°C下进行15S循环变性步骤,这也适用于富含GC的模板,但是对于低GC含量(40-45%)模板,可以将变性时间降低到5s。退火温度和时间:最佳退火温度取决于底漆序列,通常比对下的TM低2-5°C。我们建议运行温度梯度以确定最佳退火温度,另外55°C可以用作起点。取决于反应,退火时间也可以减少到5s。

Sn3N2-LiNO3-NaCl/单层BN高导热相变储能材料的热特性
热能存储引起了广泛关注,相变材料 (PCM) 因其有益的物理和化学特性而被广泛使用。虽然氮化物基盐 PCM 通常用于热能存储,但其潜热存储能力仍然有限。这项研究通过加入单层氮化硼来增强氮化物基盐用于热能存储的性能,从而提高热导率和潜热存储能力。Sn₃N₂-LiNO₃-NaCl/单层氮化硼的新型混合物具有高比热容、高潜热值和低相变温度的特点,使其成为热能存储的绝佳候选材料。在 PCM 中添加单层氮化硼可显著提高热导率,将其从 1.468 W/m·K 提高到 5.543 W/m·K。值得注意的是,这些氮化物基三元盐不会相互发生化学反应;它们的相互作用纯粹通过混合来改善热性能。该新型共混物还表现出了良好的热稳定性,在600℃时分解率仅为0.5%,熔化温度为150℃,凝固温度为130℃。三元盐的比热容达到最大值3.5 J/g·℃,表明热流速率更高,充电和放电速率也更高。复合PCM(CPCM)的储热能力在600℃时为600 kJ/kg,这些PCM的组合延长了储热时间。三元盐表现出优异的热稳定性,在100次循环中保持性能而质量没有显著减少。此外,三元盐向单层孔隙中的扩散进一步增强了其有效性。使用基于Anaconda的Jupyter Notebook和Python进行模拟分析。

DNA测序
1。底漆设计和反应设置我应该如何设计序列底漆?引物应至少长18个碱基,以确保良好的杂交尝试达到约55-60°C的T mGC含量应约为50-55%。较高的GC内容可能会导致变性不完整避免二级结构(作为发夹),尤其是在3'末端避免使用四个或更多一个基础的串链•可以避免可以混合形成二聚体的底漆不应避免使用替代性混合杂交站点。是的,重要的是考虑退火温度与底漆匹配。一般规则是,循环方案中的退火温度低于底漆的熔化温度(T m)。t m(大约)=(no a/t x 2) +(g/c x 4的否)通常,将退火温度为50-55°C,从标准引物中给出良好的序列。是否有可能减少测序预混合的量?是的,预用含有剩余的试剂。首先做出第四季度大小的反应。如果序列结果良好并且信号峰很强,则可以进一步稀释该试剂盒。始终使用套件中包含的稀释缓冲液,并将最终体积保持在20µL。保持DNA和底漆不变。请注意,仅当您使用相同类型的模板和底漆进行序列时,建议这样做。如果您使用新底漆或在DNA模板准备中进行更改,请返回到套件的第四季度稀释度并检查序列的结果。