XiaoMi-AI文件搜索系统
World File Search System纤毛

AAV介导的基因疗法减弱了Bardet-
Bardet-Biedl综合征(BBS)是一组遗传的常染色体隐性纤毛病,其特征是多种细胞类型中纤毛功能的干扰,导致肥胖,肾衰竭和失明。已知20多个因果基因,许多突变使BBSOME的功能是一种蛋白质复合物的功能,该蛋白质复合物调节货物蛋白在纤毛中的运动。BBS10基因中的突变是BBS的第二大原因,占所有病例的20%以上。最近,HSU等人的概念验证研究。2023,证明了在BBS10缺乏小鼠模型中使用小鼠BBS11的下接送递送的AAV基因治疗的潜力。在这项研究中,我们着手优化和识别带有人类BBS10基因的AAV8矢量,从而提供持续的功效和良好的临床翻译安全性。
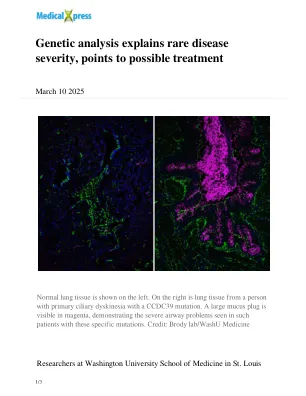
遗传分析解释了罕见疾病的严重程度,指向可能的治疗
由遗传学教授苏珊·荷兰(Susan K.除了功能失调的睫状网络之外,缺失的结构还导致某些应该具有纤毛的细胞产生粘液,这可能会导致气道问题增加。

游动和附着纤毛虫的最佳摄食
靠近水生食物链底部的纤毛微生物要么游动去寻找猎物,要么附着在基质上并产生摄食流来捕获路过的颗粒。在这里,我们使用一种流行的粘性流体球形模型来表示附着和游动的纤毛虫,其滑动表面速度可以提供纤毛流动的解析表达式。我们求解了溶解营养物浓度的平流扩散方程,其中佩克莱特数 (Pe) 反映了扩散与平流时间尺度的比率。对于固定的流体动力学功率消耗,我们问什么纤毛表面速度可以最大化微生物表面的营养通量。我们发现优化进食的表面运动取决于 Pe。对于在有限 Pe 下自由游动的微生物来说,采用“跑步机”表面运动来游动是最佳选择,但在 Pe 较大的极限下,这种跑步机解与保持生物体静止的对称偶极表面速度之间没有区别。对于附着的微生物,在 Pe 低于临界值时,跑步机解决方案是最佳的进食方式,但在 Pe 值较大时,偶极表面运动是最佳的。我们在开环数值模拟和渐近分析中验证了这些结果,并使用了基于伴生的优化方法。我们的研究结果挑战了现有的“最佳进食就是在所有佩克莱特数上最佳游动”的说法,并为海洋微生物中附着和游动解决方案的普遍性提供了新的见解。
种群中脑积水的遗传背景 -
脑积水是中枢神经系统中最常见的先天性疾病之一,经常表现出精神病合并症,特别是自闭症谱系障碍。脑积水背后的疾病机制很复杂,尚不清楚,但是已经指出了脑室和蛛网膜下腔空间中功能障碍纤毛的某些辅助。对脑积水的遗传性AETI学的更好理解,包括纤毛病的作用,可能会将见解带入潜在的共享遗传病因。在这项基于人群的病例研究中,我们首次研究了假定的脑积水候选基因的变体。使用这些数据,我们旨在研究纤毛组在脑积水中的潜在参与,并描述与自闭症谱系障碍的基因型 - 表型关联。在整个外观序列的伦敦贝克基金会综合精神病研究研究计划中筛选了一百二十一名综合基因,其中包括72名脑积水患者和4181个背景人群控制。候选基因包含感兴趣的高影响变异的基因,以系统地评估其参与睫状功能和自闭症谱系障碍。脑积水患者诊断时的中位年龄为0岁(范围0-27岁),分析年龄为22岁(11-35岁),男性为70.5%。对照年龄的中位年龄为18岁(范围11-26岁),男性为53.3%。在42例患者中鉴定出了34个基因的52个推定的脑积水相关变体(58.3%)。在脑积水病例中,我们发现高影响力蛋白改变变异的增加但不显着(优势比1.51,95%置信区间0.92-2.51,p = 0.096),这是由稀有蛋白质truncein truncating truncating truncating truncating truncating差异(频率为0.0 = 0.71 prefors p profio profiers p = 0.096)驱动的。具有高影响变体的基因中的十四个是纤毛组的一部分,而另外六个基因影响神经发生过程中CI依赖性过程。此外,在具有高影响变异的34个基因中,有15个具有蛋白质截断变体的八个基因中的3个与自闭症谱系障碍有关。由于其他疾病的症状可能被脑积水相关的症状忽略或掩盖,因此我们建议先天性脑积水患者就纤毛病和自闭症谱系疾病接受临床遗传评估。我们的结果表明,在某些情况下,脑积水作为睫状性疾病的重要性。未来在脑纤毛病中的研究可能不仅揭示了对脑积水的新见解,而且鉴于纤毛在神经发育中的重要作用,从最广泛的意义上讲,脑部疾病。

MBD5中与严重癫痫和智力残疾相关的一种新型遗传变异:对神经原发性纤毛的潜在影响
1 Coimbra大学神经科学与细胞生物学中心(CNC),3004-504 Coimbra,葡萄牙2药学学院,Coimbra大学3000-548 Coimbra,葡萄牙3,葡萄牙3研究院,3030-789 COIMBRA,PORTUGIAL 4.科尼布拉大学医学学院牙科研究所,葡萄牙6 Coimbra,葡萄牙6口腔科学创新与研究中心(CIROS),科尼布拉大学医学院,3000-075 Coimbra,Coimbra,Coimbra,Coimbra,葡萄牙7大学诊所ICAR培训中心,儿科医院,医院中心和大学至里约热内弗(CHUC),3000-602 Coimbra,Coimbra,Coimbra 9 Life Sciences,Coimbra University of Coimbra,3000-456 Coimbra,葡萄牙 *通信 *通信:jpeca@cnc.uc.cp.pt(J。) cseabra@cnc.uc.pt(c.m.s.)

在神经细胞原发性纤毛的钙动力学调节刺猬信号传导依赖性神经发生在胚胎神经管
神经干细胞增殖与神经元分化之间的平衡对于适当发展神经系统至关重要。Sonic刺猬(SHH)依次促进细胞增殖和神经表型的规范,但是负责从有丝分裂到神经源的发育转变的信号传导机制尚不清楚。在这里,我们表明,SHH通过瞬态受体电势阳离子阳离子c构件C成员3(TRPC3)(TRPC3)通过Ca 2+涌入来增强Ca 2+的活性,并通过Ca 2+涌入发育于Ca 2+涌入,并以发育阶段相互依赖的阶段相关的方式从细胞内存储中释放。这种睫状Ca 2+的活性反过来又通过下调SOX2表达和神经源性基因的上调表达来拮抗神经干细胞中的规范,增生性SHH信号,从而实现了神经元分化。这些发现表明,神经细胞睫状信号传导中的SHH-CA 2+依赖性开关触发了SHH作用从规范有限源性到神经源的开关。在该神经源信号轴上鉴定的分子机制是治疗脑肿瘤和神经发育障碍的潜在靶标。

开放式合作研究计划 - DRPM ITB
局限性:能够应用于鼻腔的制剂体积有限,包括药物从鼻粘膜的渗透性差、粘膜纤毛清除、粘液层的存在、局部酶和药物滞留时间短,这些都是阻碍通过鼻内途径吸收药物的一些因素。解决方案:鼻内制剂必须由生物相容性和无味的赋形剂组成,并避免由于粘膜纤毛清除和/或酶降解而快速消除。制剂必须具有与鼻粘膜相容的适当粘度、生理张力和 pH 值。因此,已经探索了不同的策略来克服这种给药途径的挑战。这些方法中的大多数旨在通过增加剂型在鼻粘膜中停留的时间并促进中枢神经系统药物浓度来增强分子的吸收和渗透性。4. 实现目标的策略

动物左右对称破缺策略的多样性......
摘要动物内脏器官的左右 (LR) 不对称是在胚胎发育过程中通过逐步过程建立起来的。虽然有些步骤是保留的,但动物之间采用不同的策略来启动身体对称性的破坏。在斑马鱼 (硬骨鱼类)、非洲爪蟾 (两栖动物) 和小鼠 (哺乳动物) 中,对称性破坏是由 LR 组织器处的定向流体流动引起的,这种流体流动由运动纤毛产生并被机械反应细胞感知。相比之下,鸟类和爬行动物不依赖纤毛驱动的流体流动。无脊椎动物(如蜗牛和果蝇)采用另一种不同的机制,其中对称性破坏过程由肌球蛋白和肌动蛋白分子相互作用下游获得的细胞手性支撑。在这里,我们强调了肌动球蛋白相互作用和平面细胞极性是动物之间多种 LR 对称性破坏机制的汇聚切入点。


JHEP03(2024)126
摘要:在Weyl Semimetals的磁催化场景的背景下,提出了一种在极高磁场处进行手性对称性恢复的新机制。与以前的提案相反,我们在这里表明,在非常大的磁场上,轴突场的横向速度,手性冷凝物的相模式⟨⟨⟨ψ电话,有效地变为一维及其波动破坏了该费米式冷凝物的可能的非零值。我们还表明,尽管有U(1)手性对称性未在极大的磁场上破裂,但系统的光谱由定义明确的无间隙波式激发,连接到轴轴模式,以及相关的绝缘纤毛液体与U(1)手性渗透性相关的纤毛液体。当该理论补充了动态电磁场的包含时,手性对称性再次被打破,并且可以恢复磁性催化的常规情况。