XiaoMi-AI文件搜索系统
World File Search System羟化酶

内分泌学杂志 - Bioscientifica
雄激素在斑马鱼雄性生殖发育和功能中的作用尚不清楚。为了研究这个主题,我们使用 CRISPR/Cas9 生成 cyp11c1(11 β-羟化酶)突变斑马鱼品系。我们的研究证实了最近发表的关于另一种 cyp11c1 − / − 突变斑马鱼品系的发现,并报告了 Cyp11c1 功能丧失导致的表型的新方面。我们报告称,Cyp11c1 缺陷斑马鱼主要表现出雌性第二性征,但可能拥有卵巢或睾丸。此外,我们观察到 cyp11c1 − / − 突变雄性斑马鱼严重缺乏雄激素和皮质醇。这些结果进一步证明,雄激素对于斑马鱼睾丸的形成是可有可无的,正如之前在雄激素缺乏和雄激素抗性的斑马鱼中证实的那样。在此,我们显示 cyp11c1 − / − 突变斑马鱼的睾丸表现出混乱的管状结构;并且首次证明连接睾丸和泌尿生殖口的精索管在雄激素缺乏的斑马鱼中严重发育不全。此外,我们还显示 cyp11c1 − / − 突变斑马鱼的精子发生和特征性繁殖行为受损。在 Cyp11c1 缺陷的斑马鱼的睾丸中,A 型精原细胞标记物 nanos2 的表达显著增加,而精子发生后期阶段的标记物的表达显著降低。这些观察结果表明在斑马鱼中,A 型精原细胞的产生不依赖于雄激素,但 A 型精原细胞的分化是一个依赖雄激素的过程。总体而言,我们的结果表明,虽然雄激素不是睾丸形成所必需的,但它们在决定第二性征、曲细精管的正确组织和男性生殖细胞的分化方面发挥着重要作用。

特定的NQO2抑制剂S29434,仅略微改善了多巴胺神经元在MPTP感染小鼠中的存活
摘要:多年来,有证据表明胞质喹酮还原酶NQO2在帕金森氏症诱导的多巴胺神经元变性模型中可能的贡献作用,但大多数数据已在体外获得。因此,我们问了一个问题,NQO2是否参与MPTP的体内毒性,MPTP是一种经典用于帕金森氏病诱导神经变性的神经毒素。首先,我们表明NQO2在小鼠黑质中表达,nigra多巴胺能细胞体和人多巴胺能SH-SY5Y细胞也表达。一种高度特异性的NQO2抑制剂S29434能够减少具有星形胶质细胞U373细胞的SH-SY5Y细胞的共培养系统中MPTP诱导的细胞死亡,但在SHSY5Y单一培养物中无活性。我们发现S29434仅略微防止MPTP中毒在体内中的MPTP中的黑质酪氨酸羟化酶 +细胞损失。该化合物在第7天产生了多巴胺能细胞存活的略有增加,MPTP治疗后21个,尤其是1.5 mg和3 mg/kg剂量方案。未达到统计显着性的救援效应(除了在第7天进行了一个实验),并且在最新时间点随着4.5 mg/kg剂量的降低。尽管在小鼠MPTP模型中缺乏NQO2抑制剂的强大保护活性,但我们不能排除酶在帕金森氏变性中的可能作用,尤其是因为它在多巴胺能神经元中基本上表达。

对维生素 D 关系的新见解......
2 型糖尿病 (T2DM) 是一种慢性代谢紊乱,由胰腺无法分泌足够的胰岛素或人体无法正确利用其产生的胰岛素引起 [1] 。沙特阿拉伯约有 700 万人患有糖尿病,300 多万人患有糖尿病前期,是中东地区糖尿病发病率第二高、全球第七高的国家。沙特阿拉伯的糖尿病患病率估计约为 32.8%。此外,预测 2025 年和 2030 年的患病率分别为 40.37% 和 45.36% [2] 。有家族史、年龄、肥胖和缺乏身体活动的人患 2 型糖尿病的风险会增加。曾患妊娠期糖尿病 (GDM) 的女性和糖耐量受损 (IGT) 或空腹血糖受损 (IFG) 的个人也面临患 2 型糖尿病的风险 [3] 。此外,2 型糖尿病患者更容易出现不同的并发症,包括大血管疾病(心脏病、冠状动脉疾病、中风、高血压和高脂血症)、微血管疾病(视网膜病变、神经病变和肾病)和癌症 [4] 。最近,维生素 D 缺乏症作为患 2 型糖尿病的危险因素引起了广泛关注 [5] 。根据流行病学研究,维生素 D 缺乏症影响全球多达 10 亿人 [6] 。一种重要的 25-D 羟化酶 (CYP2R1) 最近被鉴定为导致遗传性维生素 D 缺乏症的因素 [7] 。沙特人群中维生素 D 水平低的患病率很高,这被认为是由于血清 25-羟基维生素 D [25(OH) D] 的截止值过高造成的。除了营养和阳光外,各种因素都会影响维生素 D 水平。沙特人的维生素 D 缺乏症的患病率比居住在同一地区、可能食用类似饮食的非沙特人更高,这可能是因为遗传因素 [8] 。
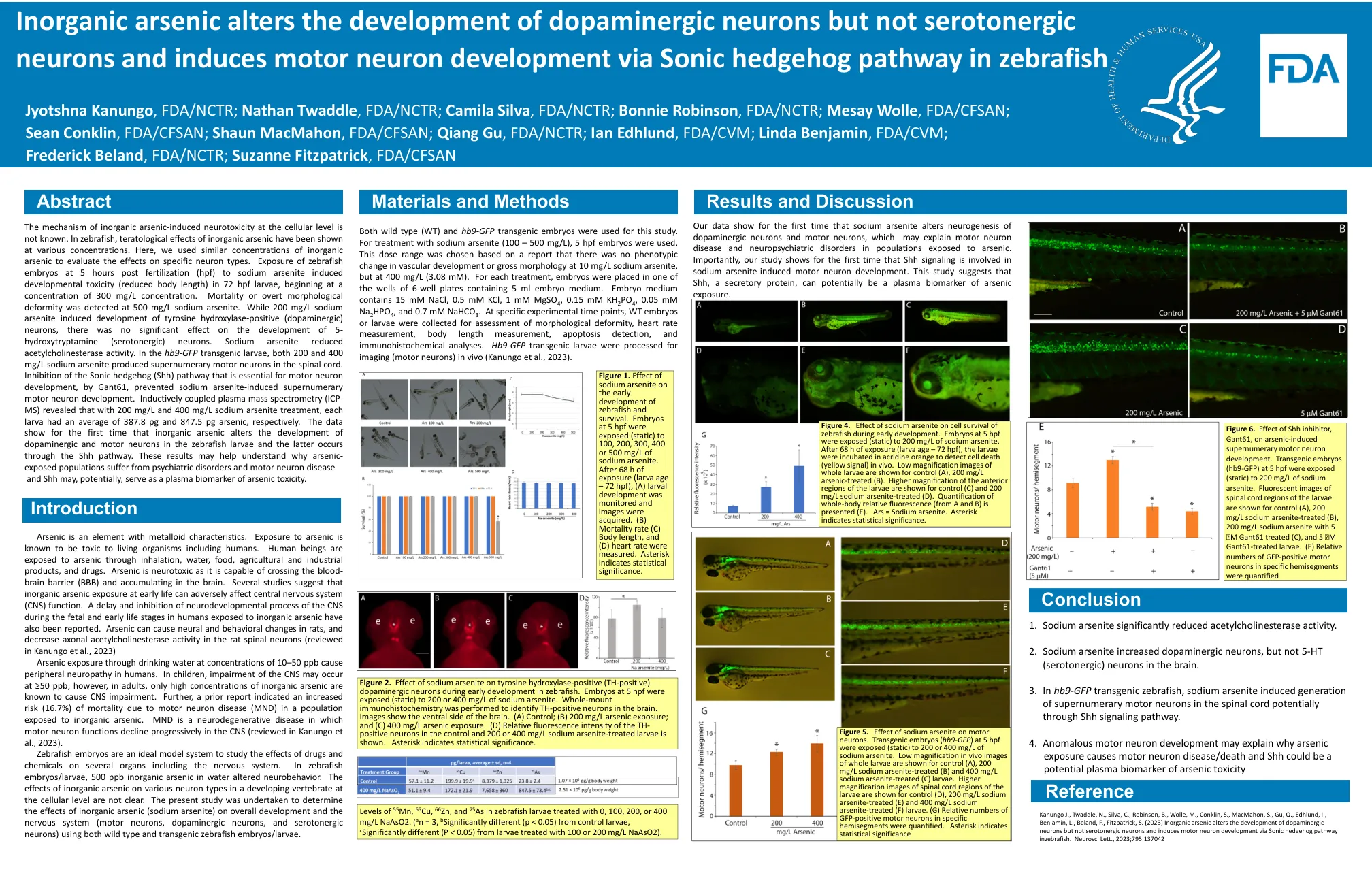
无机砷改变多巴胺能神经元的发育,但不改变血清素能神经元的发育,并通过 Sonic Hedgehog 通路诱导斑马鱼的运动神经元发育
无机砷在细胞水平上诱发神经毒性的机制尚不清楚。在斑马鱼中,不同浓度的无机砷均有致畸作用。在这里,我们使用了类似浓度的无机砷来评估其对特定神经元类型的影响。受精后 5 小时 (hpf) 的斑马鱼胚胎暴露于亚砷酸钠中,在 72 hpf 幼虫中诱发发育毒性(体长缩短),浓度从 300 mg/L 开始。在 500 mg/L 亚砷酸钠下检测到死亡或明显的形态畸形。虽然 200 mg/L 亚砷酸钠诱导酪氨酸羟化酶阳性(多巴胺能)神经元的发育,但对 5-羟色胺(血清素能)神经元的发育没有显著影响。亚砷酸钠降低了乙酰胆碱酯酶活性。在hb9-GFP转基因幼鱼中,200和400mg/L亚砷酸钠均在脊髓中产生了多余的运动神经元。通过Gant61抑制运动神经元发育所必需的Sonic Hedgehog(Shh)通路,可以阻止亚砷酸钠诱导的多余运动神经元发育。电感耦合等离子体质谱(ICP-MS)分析表明,在200mg/L和400mg/L亚砷酸钠处理下,每只幼鱼平均砷含量分别为387.8pg和847.5pg。数据首次表明无机砷改变斑马鱼幼鱼多巴胺能神经元和运动神经元的发育,后者是通过Shh通路发生的。这些结果可能有助于理解为什么接触砷的人群会患上精神疾病和运动神经元疾病,并且 Shh 可能潜在地充当砷毒性的血浆生物标志物。

着装综合症
伴有嗜酸性粒细胞增多和全身症状的药物皮疹 (DRESS) 是一种罕见、特异、危及生命的药物反应,临床表现多种多样,包括发热超过 38.5 ºC、瘙痒性斑丘疹或红斑性皮疹、血液学异常、淋巴结肿大和多器官受累。其发病率为 1/1000 至 1/10,000,估计死亡率高达 10%。迄今为止,据报道许多药物可导致 DRESS 综合征,但最常见的是抗惊厥药和磺胺类药物,尽管其发病机制尚不清楚。环氧化物羟化酶(可解毒芳香抗惊厥药的代谢物)的缺陷或不足、药物解毒不足导致可能引发免疫反应的反应性代谢物、某些 HLA 等位基因导致的易感性以及疱疹病毒的再激活,这些都表明在发病机制中发挥了作用。潜伏期从两周到六周不等。在 DRESS 综合征的临床过程中,可以看到血液、肝脏、肾脏、心脏、肺、神经、胃肠道和内分泌系统受累;以及噬血细胞综合征。DRESS 综合征的长期后遗症包括肝、肾和肾上腺功能衰竭;1 型和 2 型糖尿病、格雷夫斯病、自身免疫性溶血性贫血、狼疮、系统性硬化症和自身免疫性肠病。 DRESS 综合征的诊断很难确定,需要高度的初始临床怀疑和排除其他病因。治疗 DRESS 综合征最重要的步骤是早期诊断和及时停用致病药物。在器官受累的病例中,需要全身皮质类固醇治疗。在严重和类固醇耐药的病例中,可能需要使用更有效的免疫抑制剂或静脉注射免疫球蛋白治疗。

Monica Neugebauer 博士
摘要:酶以极高的选择性催化化学转化。通过定向进化,我们可以重新编程酶以应用于生物催化和医学。在第一部分中,我将讨论我的工作,即发现、表征和设计卤化未活化 Csp3—H 键的 FeII/α-酮戊二酸依赖性酶。我解决了一种新型赖氨酸卤化酶 (BesD) 的厌氧晶体结构,发现了能够形成九种新氯化氨基酸的同源物,并开发了酶级联以产生氯化杂环、二胺、酮酸和肽。通过结构研究和高通量筛选,我研究了该酶家族中区域选择性和催化选择性的机制基础,并利用由此获得的见解来设计羟化酶以进行卤化,其活性和选择性与天然卤化酶相当。在第二个故事中,我通过定向进化开发了新型胞嘧啶碱基编辑器 (CBE)。碱基编辑器由可编程的 DNA 结合蛋白(如催化受损的 Cas9)组成,与脱氨酶融合,可实现基因组中靶位点的精确核苷酸变化。将 C•G 碱基对转化为 T•A 的 CBE 通常比其腺嘌呤碱基编辑器 (ABE) 更大,并且具有更多不良的脱靶编辑。为了开发一类保留 ABE 有利特性的新型 CBE,我使用连续蛋白质进化来进化 ABE,以便在治疗相关位点和细胞类型内进行高效的胞嘧啶碱基编辑。这些新进化的碱基编辑器克服了现有 CBE 的几个局限性,并展示了蛋白质进化在应对生物技术挑战方面的力量。

正宗胸罩Rapa黄酮合酶1的基因编辑1产生二氢类黄酮含量的中国卷心菜
黄酮是绿色卷心菜的主要类黄酮类(Brassica Rapa subsp。pekinensis)。B. rapa基因组拥有七个黄酮醇合酶基因(BRFLS),但在功能上没有表征它们。在这里,转录组分析显示四个BRFL在中国卷心菜中主要表达。中,只有BRFLS1显示出主要的FLS活性和其他黄酮3β-羟化酶(F3H)活性,而BRFLS2和BRFLS3.1仅表现出边际F3H活性。我们使用CRISPR/CAS9介导的基因组编辑生成了BRFLS1-KNOCKOUT(BRFLS1- KO)中国卷心菜,并在T 1代中获得了没有脱靶突变的无靶向突变的无抗纯合植物,这些植物进一步前进到T 2生成t显示正常表型的T 2。UPLC-ESI-QTOF-MS分析表明,T 2植物中黄酮醇糖苷急剧降低,而二氢黄酮醇糖苷同时累积到与降低黄酮醇水平相对应的水平。定量PCR分析表明,BRFLS1-KO植物中苯基丙型和类黄酮生物合成途径的早期步骤上调。在符合BRFLS1-KO植物中,总酚类酚含量略有增强,这表明在Cabyylypopanios和Flavavonys中具有负面作用。表型调查显示,BRFLS1-KO中国卷心菜表现出正常的头部形成和生殖表型,但观察到其头部的细微形态变化。此外,与对照组相比,它们的幼苗对渗透压敏感,这表明黄酮醇在b.rapa幼苗中对渗透胁迫耐受性起积极作用。在这项研究中,我们表明CRISPR/CAS9介导的BRFLS1 -KO成功地产生了具有独特代谢性状的中国白菜的宝贵育种资源,并且CRISPR/CAS9可以有效地应用于功能性的中国白菜繁殖中。

脑损伤后的行为和转录组变化......
摘要:去甲肾上腺素 (NE) 在塑造行为结果方面起着不可或缺的作用,包括焦虑/抑郁、恐惧、学习和记忆、注意力和转移行为、睡眠-觉醒状态、疼痛和成瘾。然而,尚不清楚 NE 释放失调是这些行为适应不良取向的原因还是结果,其中许多与精神疾病有关。为了解决这个问题,我们使用了一种独特的遗传模型,其中大脑特异性囊泡单胺转运体-2 (VMAT2) 基因表达在 NE 阳性神经元中被去除,从而禁用整个大脑中的 NE 释放。我们通过将 floxed VMAT2 小鼠与在多巴胺 β -羟化酶 (DBH) 基因启动子下表达 Cre 重组酶的小鼠杂交来设计 VMAT2 基因剪接和 NE 耗竭。在这项研究中,我们对 VMAT2DBHcre KO 小鼠进行了全面的行为和转录组学表征,以评估中枢 NE 在行为调节中的作用。我们证明了 NE 耗竭会产生抗焦虑和抗抑郁样作用,改善情境恐惧记忆,改变转移行为,降低对安非他明的运动反应,并在非快速眼动 (NREM) 阶段诱导更深的睡眠。相反,NE 耗竭不会影响空间学习和记忆、工作记忆、对可卡因的反应以及睡眠-觉醒周期的结构。最后,我们使用此模型来识别在没有 NE 释放的情况下可以上调或下调的基因。我们发现突触囊泡糖蛋白 2c (SV2c) 基因表达在几个大脑区域(包括蓝斑 (LC))中上调,并且能够验证这种上调是长期社交失败脆弱性的标志。NE 系统是一个复杂且具有挑战性的系统,由于它分布在大脑中,因此涉及许多行为取向。在我们的研究中,我们揭示了 NE 神经传递在多种行为中的具体作用,并将其与分子基础联系起来,为未来理解 NE 在健康和疾病中的作用开辟了方向。
长期适应淋巴瘤细胞对缺氧的适应性是由特异性抑制剂
大量证据表明,低氧驱动恶性细胞的侵略性分子特征,而与癌症类型无关。非霍奇金淋巴瘤(NHL)是最常见的血液系统恶性肿瘤,其特征是频繁涉及多样的低氧微环境。我们研究了长期深缺氧(1%O2)对淋巴瘤细胞生物学的影响。在缺氧下≥4周,有6种测试的细胞系(RAMOS和HBL2)中只有2个。缺氧适应的(HA)B RAMOS和HBL2细胞的增殖速率降低,伴随着对氧化磷酸化和糖酵解途径的显着抑制。转录组和蛋白质组分析表明,线粒体呼吸复合物I和IV的基因和蛋白质的下调明显下调,以及线粒体核糖体蛋白。尽管观察到了糖酵解的抑制抑制,但对两个HA细胞系的蛋白质组分析表明,与葡萄糖利用的调节有关的几种蛋白质的上调,包括丙酰-4-羟化酶P4HA1的活性催化成分,这是一种重要的可药物果仁。ha细胞系显示自动/线粒体的关键调节剂的转录增加,例如神经蛋白,Bcl2相互作用蛋白3(BNIP3),BNIP3样蛋白和BNIP3 pseudogene。对缺氧的适应性进一步与凋亡失调,即Bcl2l1/bcl-XL的上调,BCl2L11/BIM的过表达,BIM与Bcl-XL的结合增加,显着提高了对A11555463的细胞对A1155463的细胞敏感性的敏感性。负责葡萄糖利用的蛋白质的上调,2。最后,在两个HA细胞系中,Akt激酶均经过过度磷酸化,并且细胞对Copanlisib的敏感性增加,这是PAN-PI3K抑制剂。总而言之,我们的数据报告有关淋巴瘤细胞适应长期缺氧的几种共享机制,包括:1。线粒体蛋白降解潜在的线粒体回收(通过线粒体)和3。增加对BCL-XL和PI3K-AKT信号的依赖性。在翻译中,抑制糖酵解,BCL-XL或PI3K-AKT级联反应可能导致靶向消除HA淋巴瘤细胞。

AAV 的候选对象和挑战
缩写:AADC,芳香族 L-氨基酸脱羧酶;AAV,腺相关病毒;ALS,肌萎缩侧索硬化症;APOE,载脂蛋白 E;ASO,反义寡核苷酸;ATXN2,共济失调蛋白 2;BBB,血脑屏障;BSCB,血脊髓屏障;CDKL5,细胞周期蛋白依赖性激酶样 5;CNS,中枢神经系统;CRISPR,成簇的规律间隔的短回文重复序列;CSF,脑脊液;DRPLA,齿状红核苍白球路易体萎缩;FTD,额颞痴呆;FUS,聚焦超声;FXTAS,脆性 X 相关震颤/共济失调综合征;GABA,γ-氨基丁酸;GAD,谷氨酸脱羧酶;GAG,糖胺聚糖; GAN,巨轴突性神经病;GBA,葡萄糖脑苷脂酶;GCH,三磷酸鸟苷环化水解酶;GDNF,胶质细胞源性神经营养因子;ICis,脑池内;ICV,脑室内;IPa,脑实质内;IT,鞘内(腰椎);IV,静脉内;LacNAc,硫酸化N-乙酰乳糖胺;MAO,单胺氧化酶;miRNA,微小RNA;MLD,异染性脑白质营养不良;MPS,粘多糖贮积症;MRgFUS,磁共振成像引导聚焦超声;MRI,磁共振成像;MSA,多系统萎缩;NCL,神经元蜡样脂褐素沉积症;NGF,神经生长因子;NTN,神经营养素;PDHD,丙酮酸脱氢酶缺乏症;Put,壳核; rAAV,重组腺相关病毒;RNAi,RNA 干扰;siRNA,短干扰 RNA,小干扰 RNA;SMA,脊髓性肌萎缩;SMARD,脊髓性肌萎缩伴呼吸窘迫;SNc,黑质致密部;SOD1,超氧化物歧化酶 1;Str,纹状体;TDP-43,TAR DNA 结合蛋白 43;TERT,端粒酶逆转录酶;TH,酪氨酸羟化酶;Th,丘脑;VTA,腹侧被盖区;ZFN,锌指核酸酶。 * 通讯作者:德克萨斯大学达拉斯分校,800 West Campbell Road, EW31, Richardson, TX 75080, USA。电子邮箱地址:Zhenpeng.Qin@utdallas.edu (Z. Qin)。