XiaoMi-AI文件搜索系统
World File Search System胚胎
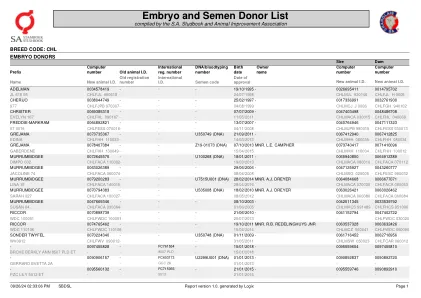
胚胎和精子捐赠者名单
10/04/2008 04/08/2006 DRIES 86 CHLFACA 030086 CHLMDK 000579 - P483/08 (DNA) PAPIE SMIT (EDMS) LTD

小鼠糖尿病诱导 电穿孔介导的基因转移至啮齿动物胚胎组织IACUC标准程序生效日期:2024年5月描述:Elec
1。如果需要进一步的长期研究,糖尿病发生后两到五天(如血糖水平所示)会接受分泌胰岛素的颗粒。 2。 胰岛素颗粒释放约0.1 U /24小时> 30天,在IACUC啮齿动物手术指南之后,使用制造商的套管卡或手术切口将皮下植入皮下植入。 如果对植入物胰岛素颗粒进行切口,请按照与渗透泵植入IACUC标准程序相同的说明,但是您不会植入渗透泵,而是植入胰岛素颗粒。 遵循胰岛素颗粒制造商提供的给药指南。 根据制造商的不同,体重不到25克的小鼠可能只需要一个胰岛素颗粒。血糖不受正常范围内(70-150 mg/dl)内的较大小鼠可能会接受两个胰岛素颗粒。 3。 应在协议中描述替代性胰岛素治疗。糖尿病发生后两到五天(如血糖水平所示)会接受分泌胰岛素的颗粒。2。胰岛素颗粒释放约0.1 U /24小时> 30天,在IACUC啮齿动物手术指南之后,使用制造商的套管卡或手术切口将皮下植入皮下植入。如果对植入物胰岛素颗粒进行切口,请按照与渗透泵植入IACUC标准程序相同的说明,但是您不会植入渗透泵,而是植入胰岛素颗粒。遵循胰岛素颗粒制造商提供的给药指南。根据制造商的不同,体重不到25克的小鼠可能只需要一个胰岛素颗粒。血糖不受正常范围内(70-150 mg/dl)内的较大小鼠可能会接受两个胰岛素颗粒。3。应在协议中描述替代性胰岛素治疗。

line1促进核分室化以抑制胚胎干细胞中的8细胞状态
。cc-by-nc-nd 4.0国际许可证(未经同行评审证明)获得的是作者/资助者,他授予Biorxiv授予Biorxiv的许可,以永久显示预印本。这是该版本的版权持有人,该版本发布于2024年9月23日。 https://doi.org/10.1101/2024.09.22.614332 doi:Biorxiv Preprint


电磁Wi-Fi辐射对鸡肉胚胎发育的影响
主要用于军事目的的无线设备领域的显着技术进步已导致一般人群的共同操作。Wi-Fi,手机和其他现代设备为其用户提供了许多优势。另一方面,它们的过度用途产生了环境负担,也称为Eleclosmog。我们当前研究的目的是观察到9天大的鸡胚胎中Wi-Fi辐射对器官组织结构的效果。在孵化的第9天,常规处理胚胎材料,以制备苏木精 - 欧洲蛋白,Picrosirius红色和周期性酸Schiff染色的组织学切片。辐射频率为2.4 GHz,平均功率密度为300 µW.m -2在整个发育中施加到第9个胚胎日,并没有从根本上影响一般的器官发生。然而,在器官的实质中,例如肝脏,脾脏,肺,肾脏和性腺以及在发育中的间充质中

胚胎发展大挑战
有关胚胎发展的研究正在进入一个新时代。作为解剖学中传统上描述性的纪律,胚胎学家已经形成了国际财团,并将重要的组织学收集数字化以保存和开放访问。胚胎发育最近在单细胞水平的颞空间转录组学上受到了更广泛的关注。这些可以预期这些跨学科的意义助长了破译胚胎发育的努力。解决其复杂性涵盖了许多挑战,这些挑战与科学,社会和政治领域相交,强调了其杰出的重要性及其固有的跨学科性质。这个领域的挑战绝不是理解复杂的生物学机制,但也具有人道主义意义。为了充分理解人类发展的机制,主要分析了胚胎发生的原则,并采用动物模型来扩大对发育过程的看法。由于最近的开拓性工作和技术进步以基于干细胞的3D方法为中心,因此我们进入了有关哺乳动物胚胎发育的历史新阶段。在脊椎动物中,现在越来越多的关注点集中在动物实验的减少。 这篇观点文章概述了这个惊人的领域的主要挑战,该挑战为基本生物医学科学以及相关的翻译方法提供了巨大的潜力,如果它们在多学科的话语中得到解决。在脊椎动物中,现在越来越多的关注点集中在动物实验的减少。这篇观点文章概述了这个惊人的领域的主要挑战,该挑战为基本生物医学科学以及相关的翻译方法提供了巨大的潜力,如果它们在多学科的话语中得到解决。

胚胎暴露于丙戊酸和新烟素会恶化发育性GABA
。cc-by-nc-nd 4.0国际许可证(未经同行评审证明)获得的是作者/资助者,他授予Biorxiv授予Biorxiv的许可,以永久显示预印本。它是制作

刺激哺乳动物胚胎发育的信号
胚胎发育受到钙(Ca 2+)信号的刺激,这些信号是通过受精的精子在卵细胞质中产生的。通过卵子形成卵。他们经过一个称为减数分裂的细胞分裂,在此过程中,它们的二倍体染色体数量减半,并通过越过新的遗传组合创建新的遗传组合。在形成过程中,卵还获得了产生Ca 2+信号所必需的细胞成分,并支持新形成的胚胎的发展。离子化钙是细胞在许多生物过程中使用的通用二信使,卵会形成“工具包”,这是信号传导所需的一组分子。 减数分裂停止了两次,这些逮捕由调节蛋白的复杂相互作用控制。 第一次减数分裂持续时间持续到青春期后,当时黄体激素激素刺激了减数分裂的恢复。 细胞周期在排卵前的第二个减数分裂分裂的中间再次停止。 男配子的结合发生在输卵管中。 配子融合后,精子从卵子的细胞内存储中释放了Ca 2+,在哺乳动物中,在哺乳动物的细胞内释放,然后是重复的Ca 2+尖峰,称为Ca 2+的振荡,在持续使用几个小时的细胞质中。 下游传感器蛋白有助于解码信号并刺激其他分子,这些分子的作用是正确发育所必需的,包括那些有助于防止其他精子细胞融合到卵中的分子,以及那些有助于从第二次减数分裂骤停,结束减数分裂并进入第一个有丝分裂细胞分裂的卵子的分子。离子化钙是细胞在许多生物过程中使用的通用二信使,卵会形成“工具包”,这是信号传导所需的一组分子。减数分裂停止了两次,这些逮捕由调节蛋白的复杂相互作用控制。第一次减数分裂持续时间持续到青春期后,当时黄体激素激素刺激了减数分裂的恢复。细胞周期在排卵前的第二个减数分裂分裂的中间再次停止。男配子的结合发生在输卵管中。配子融合后,精子从卵子的细胞内存储中释放了Ca 2+,在哺乳动物中,在哺乳动物的细胞内释放,然后是重复的Ca 2+尖峰,称为Ca 2+的振荡,在持续使用几个小时的细胞质中。下游传感器蛋白有助于解码信号并刺激其他分子,这些分子的作用是正确发育所必需的,包括那些有助于防止其他精子细胞融合到卵中的分子,以及那些有助于从第二次减数分裂骤停,结束减数分裂并进入第一个有丝分裂细胞分裂的卵子的分子。在这里我回顾了鸡蛋形成的主要步骤,讨论生成Ca 2+

鉴定多能胚胎干细胞生物学的钥匙细胞 - 细胞相互作用
上下文细胞间交流对于多细胞生物的发展和维护至关重要。最近的研究强调了哺乳动物胚胎中胚外细胞(滋养剂和低纤维细胞)和多能胚胎细胞(epiblast)之间通信的重要性[1,2,3]。具体而言,由小细胞产生的细胞外基质在控制多脂蛋白层状干细胞的细胞增殖中起关键作用。我们对猪胚胎干细胞的研究已经证实了细胞外基质在影响调节层细胞多能性的信号通路和转录因子中的重要性[4]。我们最原始的观察结果之一依赖于四叠蛋白CD9,CD81和CD63表达的检测主要是在滋养剂中(未发表)。这些四翼烷蛋白是细胞外囊泡(EV)的已知标记,它们是各种细胞类型的小脂质囊泡[5]。evs参与将信号蛋白,细胞因子和转录因子转运到受体细胞,影响各种过程,包括免疫反应,肿瘤进展和胚胎发育[6,7]。尽管已经在小鼠胚胎中研究了胚胎和胚外细胞之间的EV介导的细胞对细胞通信,但焦点主要是由小鼠多能干细胞分泌的电动汽车[8,9]。这项研究旨在研究EVS在促进交流和影响这些细胞类型的生物学的促进性滋养剂细胞和多能细胞分泌的电动汽车的作用。这项研究由ANR STEM4PIGS(2025-2028)资助,并将支持从2025年9月开始的博士生。

hridaya的阿育吠陀概念及其胚胎学...
根据阿育吠陀(Ayurveda)的说法,心脏或hridaya是人体必不可少的器官之一。对Hridaya造成的任何伤害都可能导致严重的疾病,严重的问题甚至死亡,因为它是Trimarams和Dasha Pranaayatana之一。根据阿育吠陀文本,心脏是具有血泵外具有广泛功能的器官。根据阿育吠陀经典,hridaya是智力和生活的所在地,并控制着身体,思想和感官。rakta和kapha的本质构成了心。因此,被杀死的Rakta和Kapha对于理解许多心脏病的病理生理学至关重要。在本文中,我们主要着重于Hridaya Nirukti,Hridaya Uttpatti,以及与阿育吠陀观点中胚胎学发展中结构畸形有关的疾病。