XiaoMi-AI文件搜索系统
World File Search System苏氨酸
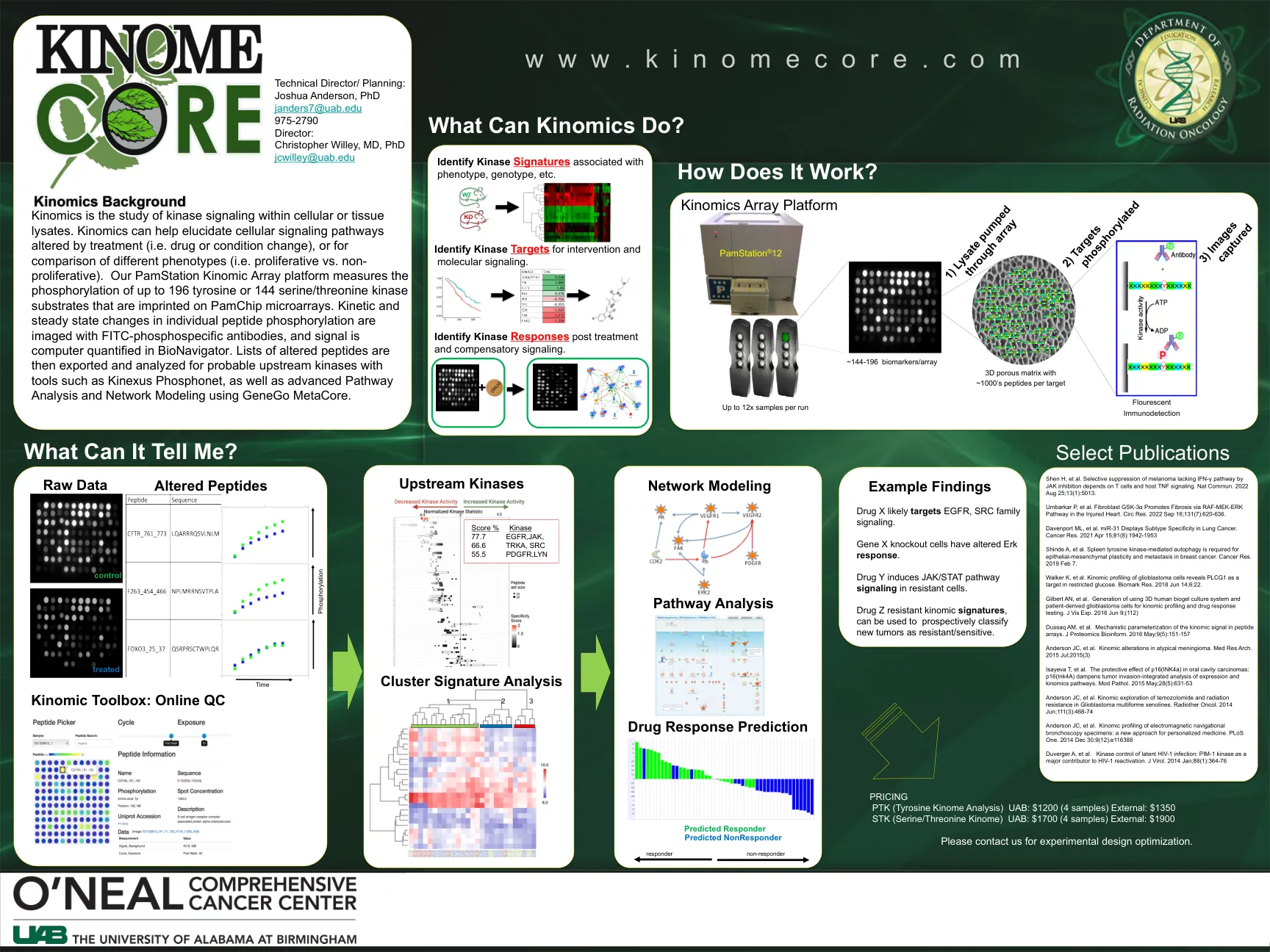
kinomics数组平台
激化组是对细胞或组织裂解物中激酶信号传导的研究。激素学可以帮助阐明因治疗而改变的细胞信号传导途径(即药物或状况变化),或用于比较不同的表型(即增殖与非增生性)。我们的pamstation kinomic阵列平台测量了最多196个酪氨酸或144个丝氨酸/苏氨酸激酶底物的磷酸化,这些丝氨酸/苏氨酸激酶底物印在Pamchip微阵列上。动力学和稳态的单个肽磷酸化的变化是用FITC磷酸化抗体成像的,并且信号在Bionavigator中进行了定量。然后,将改变肽的改变的肽列表通过使用Kinexus phosphonet等工具,以及使用Genego Metacore的高级途径分析和网络建模来输出并分析可能的上游激酶。

RIPK1和RIPK3抑制剂:针对炎症的潜在武器治疗糖尿病并发症
糖尿病是一种代谢疾病,其特征是由于多种病因因素而引起的慢性高血糖。长期代谢应激会诱导有害的炎症,导致慢性并发症,主要是糖尿病眼科,糖尿病性心血管并发症和糖尿病性肾病。由于糖尿病并发症是残疾和死亡的主要原因之一,在组合疗法中使用抗肿瘤的糖尿病正在增加。对靶向炎症途径的显着调节剂的兴趣越来越大,特别是受体互动丝氨酸/苏氨酸激酶-1(RIPK1)(RIPK1)和受体互动丝氨酸/苏氨酸 - 激酶-3(RIPK3)(RIPK3),作为用于治疗偶然性疾病的药物靶标。在这篇综述中,我们旨在提供有关RIPK1和RIPK3的作用和药物开发机制的最新摘要,这些研究在慢性炎症和免疫力中至关重要,这些研究与糖尿病并发症有关,与糖尿病并发症有关,这些并发可能是对选择性RIPK1和RIPK1的潜在的,并且是RIPK1的潜在,并且并发症。


运动在 2 型糖尿病管理中的病理生理学和作用机制
肥胖等病理状况会伴有游离脂肪酸水平的升高以及白细胞介素 6 (IL-6) 和肿瘤坏死因子 α (TNF α) 等炎性细胞因子的升高,这些因子与肝脏和骨骼肌水平的胰岛素抵抗有关,通过促进丝氨酸和苏氨酸 (Ser/Thr 激酶) 的磷酸化而不是 IRS 等蛋白质的酪氨酸的磷酸化 ( 21 )。虽然导致骨骼肌 IR 的确切机制尚不完全清楚,但最重要的理论之一与细胞内脂肪和脂肪酸代谢物的升高有关,这种现象称为脂毒性 ( 22 )。
一种新颖的发现:1961年的Poly-U Matthaei-Nirenberg实验不能充当mRNA
表明,“ poly-u刺激了许多其他许多其他氨基酸纳入蛋白质,例如亮氨酸,异亮氨酸,苏胺,苏氨酸,精氨酸,精氨酸,组氨酸,赖氨酸,丝氨酸,色氨酸和脯氨酸””由poly-u刺激,也不知道为什么马特塞伊(Matthaei)和尼伦贝格(Nirenberg与苯丙氨酸相对应的聚-U刺激”意味着“总4个碱基的特异性”仅对应“总4种4种氨基酸”,而不是“总共20种氨基酸”。“特殊性”的概念是一个理论上的错误)。

DCC-3084 是一种 RAF 二聚体抑制剂,可广泛抑制 BRAF I、II、III 类、BRAF 融合和 RAS 驱动的实体瘤,从而导致
ARAF,丝氨酸/苏氨酸蛋白激酶 A–快速加速纤维肉瘤;ATP,三磷酸腺苷;AUC,浓度时间曲线下面积;AUC 0–last,从时间 0 到最后测量浓度的 AUC;BCRP,乳腺癌耐药蛋白转运蛋白;BID,每日两次;BRAF,v-Raf 鼠肉瘤病毒致癌基因同源物 B1;CNS,中枢神经系统;CRAF,丝氨酸/苏氨酸蛋白激酶 C-Raf;CSF,脑脊液;DFG,天冬氨酸-苯丙氨酸-甘氨酸;DMSO,二甲基亚砜;ELISA,酶联免疫吸附试验;ERK,细胞外信号调节激酶;GTP,三磷酸鸟苷;hrs,小时;IC 50,半数最大抑制浓度; Kp uu,非结合分配系数(游离脑浓度/游离血浆浓度);KRAS,Kirsten RAS;M,摩尔;MDR1,多药耐药突变转运体;MEK,丝裂原活化蛋白激酶激酶;NRAS,神经母细胞瘤 RAS;PERK,蛋白激酶 R 样内质网激酶;PK,药代动力学;po,口服;pRSK,磷酸化 RSK;QD,每日一次;RAF,快速加速性纤维肉瘤;RAS,大鼠肉瘤小 GTPase 蛋白;RSK,核糖体 s6 激酶;SEM,均值标准误差;t 1/2,半衰期;TGI,肿瘤生长抑制;T. sol,热力学溶解度;WT,野生型。

基于喹唑啉的 VEGFR-2 抑制剂作为潜在的抗...
ROS 活性氧 RPTEC 肾近端小管上皮细胞 SAR 构效关系 Sck 血清肌酸激酶 Src 肉瘤 TGI 肿瘤生长抑制 Thr 苏氨酸 Tie-2 血管生成素-1 受体 TSP 血小板反应蛋白 Tyr 酪氨酸 Val 缬氨酸 VEGF 血管内皮生长因子 VEGF-A 血管内皮生长因子 A VEGF-B 血管内皮生长因子 B VEGF-C 血管内皮生长因子 C VEGF-D 血管内皮生长因子 D VEGF-E 血管内皮生长因子 E VEGF-F 血管内皮生长因子 F

药物成瘾的分子神经生物学
偶联因子(称为 G 蛋白)、第二信使 [例如 cAMP、cGMP、Ca 2 +、一氧化氮 (NO) 和磷脂酰肌醇 (PI) 和花生四烯酸 (AA) 的代谢物] 和蛋白质磷酸化(包括蛋白激酶对磷蛋白的磷酸化和蛋白磷酸酶对磷蛋白的去磷酸化),介导神经递质对其靶神经元的多种作用。第二信使依赖性蛋白激酶(例如由 cAMP 或 Ca 2 + 激活的蛋白激酶)被归类为蛋白丝氨酸/苏氨酸激酶,因为它们在丝氨酸或苏氨酸残基上磷酸化底物蛋白。每个第二信使依赖性蛋白激酶磷酸化都引用一组特定的底物蛋白(可视为第三信使),从而导致神经递质的多种生物反应。对神经递质的多种生物反应可分为三大类。在某些情况下,细胞内信使介导某些神经递质在打开或抑制特定离子通道方面的作用。然而,细胞内信使介导神经递质对其目标神经元的许多其他作用。有些相对短暂,涉及调节神经元的一般代谢状态、合成或释放神经递质的能力以及各种受体和离子通道对各种突触输入的功能敏感性。其他相对长寿,通过调节目标神经元中的基因表达来实现。因此,神经递质通过调节细胞内信使通路和改变基因转录和蛋白质合成,改变了靶神经元中受体和离子通道的数量和类型、这些神经元中细胞内信使系统的功能活动,甚至改变了神经元形成的突触的形状和数量。该图是为了说明细胞内信使系统可以放大神经递质的作用:神经递质与其受体(第一信使水平)结合的单一事件可以通过第二、第三、第四等信使水平起作用,从而产生越来越广泛的生理效应。改编自 Hyman 和 Nestler 1993。
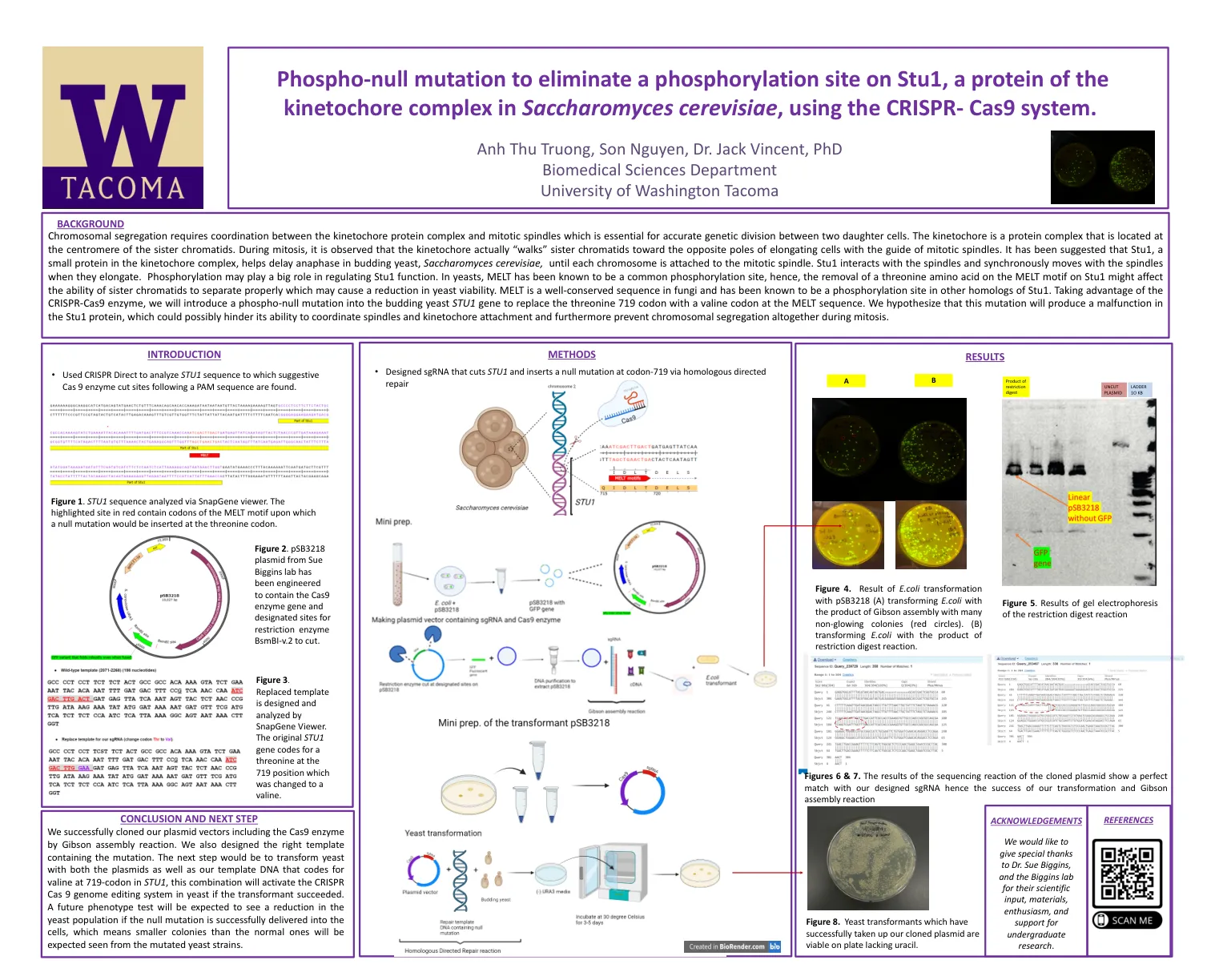
利用 CRISPR 技术进行磷酸化无效突变,消除酿酒酵母动粒复合体蛋白 Stu1 上的磷酸化位点
染色体分离需要动粒蛋白复合物和有丝分裂纺锤体的协调,这对于两个子细胞之间的准确遗传分裂至关重要。动粒是一种位于姊妹染色单体着丝粒的蛋白复合物。在有丝分裂过程中,可以观察到动粒实际上是在有丝分裂纺锤体的引导下将姊妹染色单体“引导”到伸长细胞的相反极点。有人提出,动粒复合物中的小蛋白 Stu1 有助于延迟芽殖酵母酿酒酵母的后期,直到每条染色体都附着在有丝分裂纺锤体上。Stu1 与纺锤体相互作用,并在纺锤体伸长时与其同步移动。磷酸化可能在调节 Stu1 功能方面发挥重要作用。在酵母中,MELT 是一种常见的磷酸化位点,因此,去除 Stu1 上 MELT 基序上的苏氨酸氨基酸可能会影响姐妹染色单体正确分离的能力,从而导致酵母活力下降。MELT 是真菌中保存良好的序列,并且已知是 Stu1 其他同源物中的磷酸化位点。利用 CRISPR-Cas9 酶,我们将在芽殖酵母 STU1 基因中引入磷酸化无效突变,以将 MELT 序列中的苏氨酸 719 密码子替换为缬氨酸密码子。我们假设这种突变会导致 Stu1 蛋白发生故障,这可能会阻碍其协调纺锤体和着丝粒附着的能力,并进一步阻止有丝分裂期间染色体分离。

现在就做患者需要的
1风险定义根据所使用的准则和工具而有所不同:使用SEER数据的内部估计诊断时阶段; ET =内分泌疗法; PIK3CA-MUT =磷脂酰肌醇3-激酶,催化,α多肽突变; HR+=激素受体阳性; ER+=雌激素受体阳性; HER2 =人表皮生长因子受体2; E/MBC =早期/转移性乳腺癌; CDKI =细胞周期蛋白依赖性激酶抑制剂; TKI =酪氨酸激酶抑制剂; neoadj = neoadjuvant; adj =辅助; ADC =抗体药物结合; bev = bevacizumab; BIC =一流的; SOC =护理标准; Akt =丝氨酸/苏氨酸激酶1; mtor =雷帕霉素的哺乳动物靶标