XiaoMi-AI文件搜索系统
World File Search System转座

DNA转座子通过富集转录因子结合基序
从基因组的非编码区域通过突变依次出现。除其他外,此类突变分析转录并创建一个新的开放阅读框(ORF)。尽管ORF出现的机制有充分的文献证明,但对实现新转录事件的机制知之甚少。然而,在许多物种中,已经报道了基因组所有区域的缺乏和非常突出的转录之间的连续体。在这项研究中,我们使用新组装的基因组和七个果蝇的近交系列的转录组和转录组搜索了从头转录本,该基因组和一个来自六个欧洲和一个非洲人口的近交系列。此设置使我们能够检测Sam ple特定的从头转录本,并将其与其他样品中的同源非转录区以及遗传和基因间控制序列进行比较。我们研究了与转换元件(TES)的关联,并富集了从头开始出现的转录本上游的转录因子基序,并将其与调节元素进行了比较。我们发现,从头的成绩单与TES重叠的频率比偶然性的频率更高。新转录本的出现cor与高鸟嘌呤 - 环蛋白含量和TE表达的区域有关。此外,从头转录本的上游区域高度丰富了调节基序。这种基序在与TES(尤其是DNA TES)重叠的新转录物中更丰富,并且比上游的“非转录同源物”更保守上游。总体而言,我们的研究表明,TE插入对于转录本的出现很重要,部分是通过引入DNA te家族的新调节图案。

生物化学年度评论 利用 CRISPR 相关 Tn7 样转座子进行天然和人工引导 RNA 定向转座
CRISPR-Cas(成簇的规律间隔的短回文重复序列 - CRISPR 相关核酸酶)防御系统已多次自然地用于指导 RNA 定向转座。在所有情况下,转座子 Tn7 相关的各种元件都参与了转座。Tn7 严格控制转座;只有当专用靶位选择蛋白识别特殊靶标时,转座酶才会被激活。Tn7 和与 CRISPR-Cas 系统合作的 Tn7 样元件进化出了互补的靶向途径:一条途径识别染色体中高度保守的位点,另一条途径靶向能够进行细胞间转移的移动质粒。Tn7 和 Tn7 样元件将单一整合传递到它们识别的位点,并控制整合事件的方向,为未来用作可编程基因整合工具提供了潜力。早期研究表明,引导 RNA 介导的转座系统可以适应不同的宿主,甚至在微生物群落内,这表明将这些系统设计为强大的基因编辑工具具有巨大的潜力。

R2逆转座子N末端结构域的保守和不同的DNA识别特异性和功能
R2非长末端重复(非LTR)逆转录子是多细胞真核生物中分布最广泛的移动遗传元件之一,并且对在人类基因组的转基因补充中的应用显示出希望。他们以精致的特异性将新基因插入28S核糖体DNA中的保守位点。r2进化枝是由逆转录子编码的蛋白的N末端的锌指(ZF)数量定义的,该蛋白被认为是为添加赋予DNA位点特异性的。在这里,我们阐明了进化枝之间的R2 N末端结构域的DNA识别的一般原则,并具有广泛的,具体的识别,仅需要一个或两个紧凑型域。DNA结合和保护测定法证明了广泛共享以及进化枝特异性的DNA相互作用。基因插入测定识别足以用于目标位点插入的N末端结构域,并揭示了进化枝特异性ZFS中第二链裂解或合成中的作用。我们的结果对理解非LTR逆转录座插入机制的进化多样化以及基于逆转录座子的基因疗法的设计具有意义。

Tagify™ i5 UMI 适配器加载转座酶 24/96 ...
简介 seqWell 的 Tagify™ i5 UMI 适配器负载转座酶试剂旨在催化通过 Tn5 转座酶用寡核苷酸有效载荷片段化和标记 DNA 的反应。具体而言,这些试剂可提供由全长、与 Illumina 兼容的 P5/i5/UMI/R1 引发序列组成的寡核苷酸,这些序列还包含 10 个碱基的条形码和 10 个碱基的唯一分子标识符 (UMI) 区域。这些试剂可作为靶向测序检测的一部分加入,例如 UDiTaS 1 或 RGen-Seq 2 应用、CRISPR QC 以及细胞和基因工程 QC。该产品以 24 种或 96 种不同的条形码 UMI 试剂形式提供。本用户指南介绍了试剂的一般用途,并非旨在作为特定文库制备方法的完整协议。建议个人用户查看其应用程序 3 并根据需要进行修改。

基于转座子的汽车T细胞疗法的系统
改进的SB Transposon平台Magnani CF(2016)Oncotarget 7(32):51581-51597;EP20140192371“改进了产生转基因细胞的方法”; Turazzi(2018)Br J Haematol 182(6):939-943; Magnani CF(2018)Hum gen ther 29(5):602-613; Rotiroti MC等人,在印刷中,分子治疗2020

可转座元件在正常和白血病造血中的茎状Giacomo Grillo 1,&,Bettina Nadorp 1,&,Aditi Qamra 1,Amanda Mitchell 1
1。玛格丽特癌症中心,大学健康网络,多伦多,加拿大安大略省,M5G 1L7 2。 多伦多大学多伦多大学医学生物物理学系,加拿大安大略省,M5G 1L7 3。 医学肿瘤学和血液学系,医学系,大学卫生网络,多伦多,加拿大安大略省多伦多,M5G 2M9 4。 多伦多大学多伦多大学医学系,加拿大安大略省,M5G 1A1 5。 BARTS癌症研究所,伦敦皇后大学的Barts癌症研究所,英国伦敦伦敦伦敦广场,EC1M 6BQ 6。 伦敦玛丽玛丽大学表观遗传学中心,英国伦敦,E1 4NS 7。 多伦多大学多伦多分子遗传学系,加拿大安大略省,M5S 1A8 8。 安大略省癌症研究所,多伦多,安大略省,加拿大,M5G 0A3&。 同等贡献玛格丽特癌症中心,大学健康网络,多伦多,加拿大安大略省,M5G 1L7 2。多伦多大学多伦多大学医学生物物理学系,加拿大安大略省,M5G 1L7 3。医学肿瘤学和血液学系,医学系,大学卫生网络,多伦多,加拿大安大略省多伦多,M5G 2M9 4。多伦多大学多伦多大学医学系,加拿大安大略省,M5G 1A1 5。 BARTS癌症研究所,伦敦皇后大学的Barts癌症研究所,英国伦敦伦敦伦敦广场,EC1M 6BQ 6。 伦敦玛丽玛丽大学表观遗传学中心,英国伦敦,E1 4NS 7。 多伦多大学多伦多分子遗传学系,加拿大安大略省,M5S 1A8 8。 安大略省癌症研究所,多伦多,安大略省,加拿大,M5G 0A3&。 同等贡献多伦多大学多伦多大学医学系,加拿大安大略省,M5G 1A1 5。BARTS癌症研究所,伦敦皇后大学的Barts癌症研究所,英国伦敦伦敦伦敦广场,EC1M 6BQ 6。 伦敦玛丽玛丽大学表观遗传学中心,英国伦敦,E1 4NS 7。 多伦多大学多伦多分子遗传学系,加拿大安大略省,M5S 1A8 8。 安大略省癌症研究所,多伦多,安大略省,加拿大,M5G 0A3&。 同等贡献BARTS癌症研究所,伦敦皇后大学的Barts癌症研究所,英国伦敦伦敦伦敦广场,EC1M 6BQ 6。伦敦玛丽玛丽大学表观遗传学中心,英国伦敦,E1 4NS 7。多伦多大学多伦多分子遗传学系,加拿大安大略省,M5S 1A8 8。安大略省癌症研究所,多伦多,安大略省,加拿大,M5G 0A3&。 同等贡献安大略省癌症研究所,多伦多,安大略省,加拿大,M5G 0A3&。同等贡献

核糖体 DNA 与转座因子之间的动态相互作用:基因组学和细胞遗传学的视角
尽管核糖体 DNA 和转座因子都是基因组的显著特征,但乍一看,它们都是没有太多共同点的遗传因子:核糖体 DNA 主要被视为管家基因,支持所有主要基因组功能,而转座因子通常被描绘成自私和破坏性的。这些对立的特征也反映在其他属性中:串联组织(核糖体 DNA)与分散组织(转座因子);协同进化(核糖体 DNA)与多样化进化(转座因子);延长基因组稳定性的活动(核糖体 DNA)与缩短基因组稳定性的活动(转座因子)。回顾已报道的核糖体 DNA-转座因子相互作用的相关实例,我们注意到两种重复类型至少具有四个结构和功能特征:(1)它们是在进化时间尺度上塑造基因组的重复 DNA,(2)它们交换结构基序并可以进入共同进化过程,(3)它们是严格控制的基因组应激传感器,在衰老/老化中发挥关键作用,以及(4)它们具有共同的表观遗传标记,例如 DNA 甲基化和组蛋白修饰。在这里,我们概述了核糖体 DNA 和转座因子的结构、功能和进化特征,讨论了它们的作用和相互作用,并强调了我们在理解核糖体 DNA-转座因子关联方面的趋势和未来方向。

转座元素插入与绿色蝴蝶hypolimnas misippus
1剑桥大学,剑桥大学,剑桥CB2 CB2 3EJ,英国2号生命之树计划,惠康桑格研究所,英国欣克斯顿3号弗里德里希·米舍(Max Planck Society of Max Planck Society of Max Planck Society tübingen,德国),德国,德国4号,4 4号,卢比根4号,卢旺达,卢旺达5 MPAL 5 MPAL 5 MPAL,RWANDA CENTIPLE,NANDA肯尼亚,莱基皮亚6日生物科学学院,加的夫大学,加的夫CF 10 3AX,英国7英国生态与水文学中心,Wallingford OX10 8BB,英国8 InstitutBotànicede Barcelona(IBB)(IBB),CSIC-CMCNB,CSIC-CMCNB,BARCELONA,BARCELORA康沃尔郡,佩林TR10 9FE,英国11 Turkana Basin Institute,Stony Brook University,Stony Brook,NY 11794,美国12,美国爱丁堡大学进化生物学研究所,英国爱丁堡大学
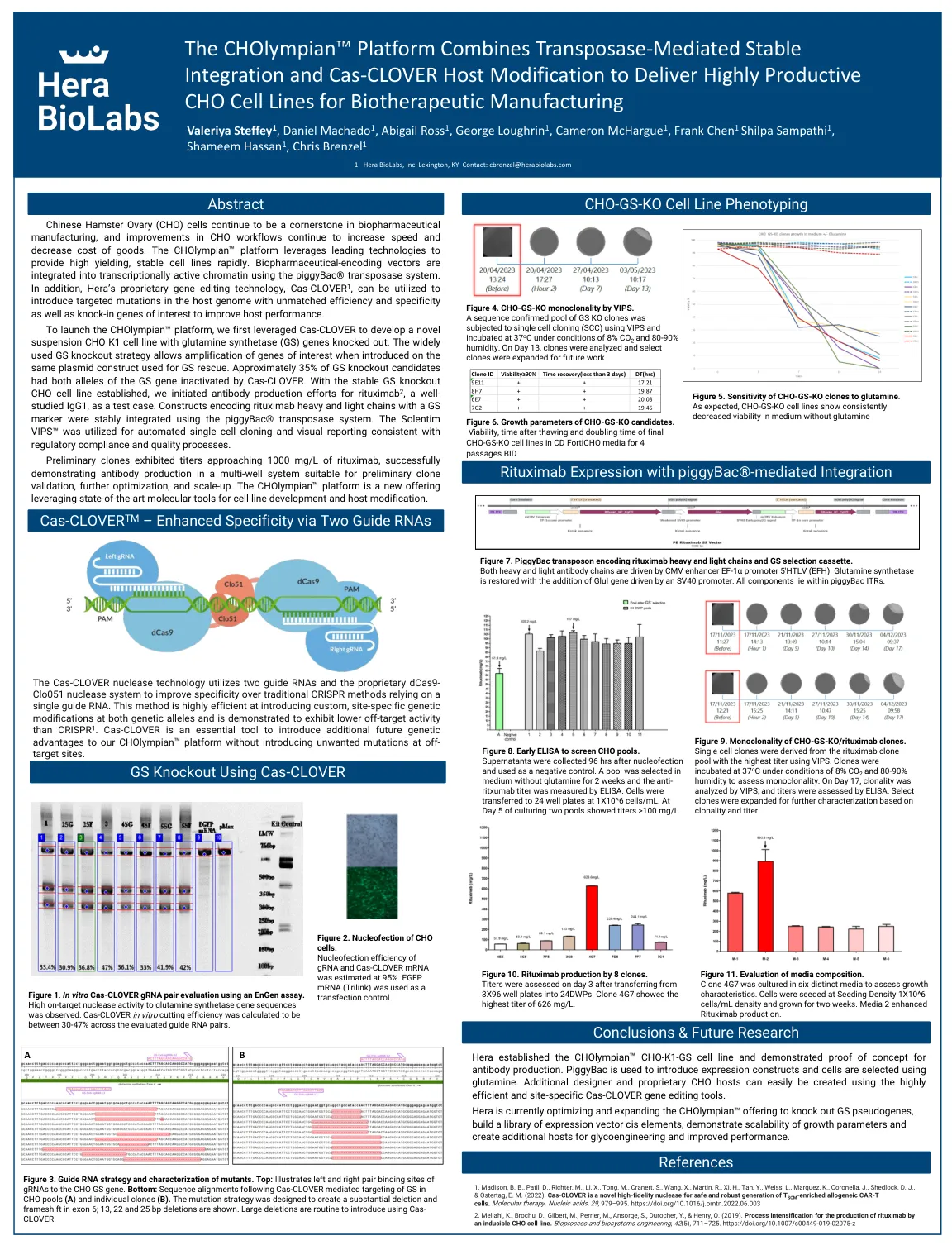
CHOlympian 平台结合转座酶...
平台,我们首先利用 Cas-CLOVER 开发了一种敲除谷氨酰胺合成酶 (GS) 基因的新型悬浮 CHO K1 细胞系。广泛使用的 GS 敲除策略允许在用于 GS 拯救的相同质粒构建体上引入目的基因时对其进行扩增。大约 35% 的 GS 敲除候选细胞的两个等位基因都被 Cas-CLOVER 灭活。在建立稳定的 GS 敲除 CHO 细胞系后,我们开始针对利妥昔单抗 2(一种研究充分的 IgG1)进行抗体生产,作为测试案例。使用 piggyBac® 转座酶系统稳定整合了编码具有 GS 标记的利妥昔单抗重链和轻链的构建体。Solentim VIPS

通过 RNA 引导转座对 Anabaena sp. PCC 7120 进行基因组工程改造
摘要:在基因组工程中,传入 DNA 的整合依赖于分裂细胞产生的酶,这一直是提高 DNA 插入频率和准确性的瓶颈。最近,据报道,使用 CRISPR 相关转座酶 (CAST) 的 RNA 引导转座在大肠杆菌中非常有效且具有特异性。在这里,我们开发了 Golden Gate 载体来在丝状蓝藻中测试 CAST,并证明它在鱼腥藻属菌株 PCC 7120 中有效。含有 CAST 和工程转座子的相对较大的质粒通过使用自杀或复制质粒的结合成功转移到鱼腥藻中。编码靶标前导链但不编码反向补体链的单向导 (sg) RNA 与 sgRNA 中包含的原间隔子相关基序 (PAM) 序列有效。在对两个不同靶位点进行分析的六种病例中,有四种的插入位点位于 PAM 之后正好 63 个碱基处。复制质粒上的 CAST 具有毒性,可用于治愈质粒。在分析的所有六种情况下,只有由从左到右元素的序列定义的转座子货物被插入目标位点;因此,RNA 引导的转座是由剪切和粘贴引起的。没有内源转座子通过暴露于 CAST 酶而重新动员。这项工作为通过 RNA 引导的转座在丝状蓝藻中进行基因组编辑奠定了基础,无论是在培养中还是在复杂群落中。关键词:鱼腥藻、CRISPR 相关转座子 (CAST)、基因组工程、RNA 引导的转座、minion 测序、从头基因组组装 ■ 简介