XiaoMi-AI文件搜索系统
World File Search System造血

儿科肿瘤学和造血细胞移植的流行病学从2001 - 2019年开始送往美国重症监护病房
近年来,随着实时流媒体技术的快速发展,电子商务实时流已经成为一种重要的消费者购物体验形式(Luo X.等,2024; Luo L.等,2024)。同时,人工智能技术的进步导致了企业对虚拟流的广泛采用,因为它们的优势,例如成本效益,高生产率和24/7的可用性。然而,尽管面临着巨大的市场机会,但虚拟流媒体也遇到了挑战,包括缺乏社会存在和简短的消费者互动时间(Gao等,2024)。为了增强互动性并在现场流媒体会话中创造了更具吸引力的氛围,虚拟流媒体已经开始模仿人类流媒体的语言和行为,尤其是通过采用亲切的昵称来与消费者建立情感联系(Leech,2014; Wang,2022; Cheng,202222)。尽管做出了模仿人类互动方式的努力,但深情的昵称在人与光明关系中的影响是否类似于人际关系中的人际关系,这是进一步探索的关键问题。这个问题不仅对指导虚拟流媒体的沟通策略具有重要的实际含义,还增强了我们对消费者与服务机器人之间的人类关系的理解。
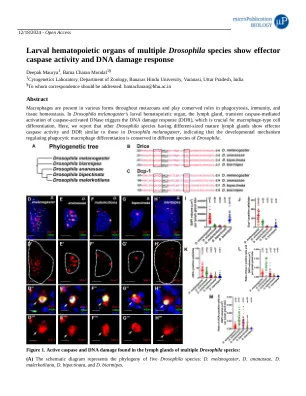
多种果蝇幼虫造血器官表现出效应胱天蛋白酶活性和DNA损伤反应
1882 年,埃利·梅契尼科夫 (Élie Metchnikoff) 在海星幼虫中发现了巨噬细胞,这种细胞通过吞噬外来物质来破坏外来物质。他将这一过程描述为吞噬作用 (Underhill 等人,2016)。后续研究表明,巨噬细胞在整个后生动物中都得到了保留,在调节发育、组织修复、体内平衡和先天免疫方面表现出额外的功能 (Lazarov 等人,2023;Park 等人,2022)。在三胚层动物中,吞噬细胞由于开放的循环系统而穿过体腔并清除细胞碎片或病原体 (Maheshwari,2022;Banerjee 等人,2019)。在哺乳动物中,常驻组织巨噬细胞在早期胚胎阶段从卵黄囊和红细胞-髓系前体细胞发育而来,并在整个生命过程中具有自我更新能力。单核细胞衍生的巨噬细胞也与快速补充的组织有关,例如肠道(Lazarov 等人,2023;Lee & Ginhoux,2022;Park 等人,2022)。在从单细胞生物进化到高度复杂的脊椎动物的过程中,巨噬细胞的作用和吞噬过程在很大程度上保持了下来(Yutin 等人,2009)。然而,吞噬巨噬细胞分化的潜在机制仍不清楚。


造血干细胞移植患者的全肠胃外营养总营养的临床影响
Results: Our results showed that total parenteral nutrition (TPN) effectively mitigated weight loss on days 10 and 14 and the day before discharge, while also improving albumin (33.41 ± 4.57 in the control group, 34.87 ± 4.08 in the TPN group, p < 0.05; 33.72 ± 3.52 in the control group, 35.27 ± 4.04 in the TPN group, p < 0.05; 34.09 ± 4.44 in the control group, 35.55 ± 3.87 in the TPN group, p < 0.05) and prealbumin (245.18 ± 79.94 in the control group, 274.26 ± 86.73 in the TPN group, p < 0.05; 233.27 ± 79.57 in the control group, 279.34 ± 80.20 in the TPN group, p < 0.01;对照组的247.24±83.29,在同一时期,TPN组为280.65±100.22,p <0.05)。此外,非TPN和TPN组之间的CRBSI发生率或肝功能没有显着差异。此外,TPN组的住院时间较短(对照组为48.06±13.90,TPN组的42.13±14.22*,p <0.05),意外重新携带的发生率较低,对照组为37.1%(在对照组中为37.1%,在TPN组为21.4%,TPN组,P <0.05),P <0.05)。

造血干细胞移植期间的营养(...
1尽可能使用单独打包或包裹的部分。2仅允许3年以上,并避免在孩子面前打开包装。3货架稳定性是指在室温下打开之前可以存储的未打开的罐装,瓶装或包装食品;打开后可能需要制冷。4在打开前用水洗罐头。5年龄的儿童允许5个巴氏消毒的蜂蜜产品,并在移植后九个月以上。6草药和草药补充剂不接受纯度或对健康的影响,也可能包含有害成分。7在最低温度为70°C的30分钟的最低温度下烘烤干果,将使水果消灭任何昆虫及其卵。

造血细胞移植治疗血液病...
AML,急性髓细胞白血病;ALL,急性淋巴细胞白血病;CLL,慢性淋巴细胞白血病;CML,慢性髓细胞白血病;DLBCL,弥漫性大B细胞淋巴瘤;MDS,骨髓增生异常综合征;MF,骨髓纤维化..
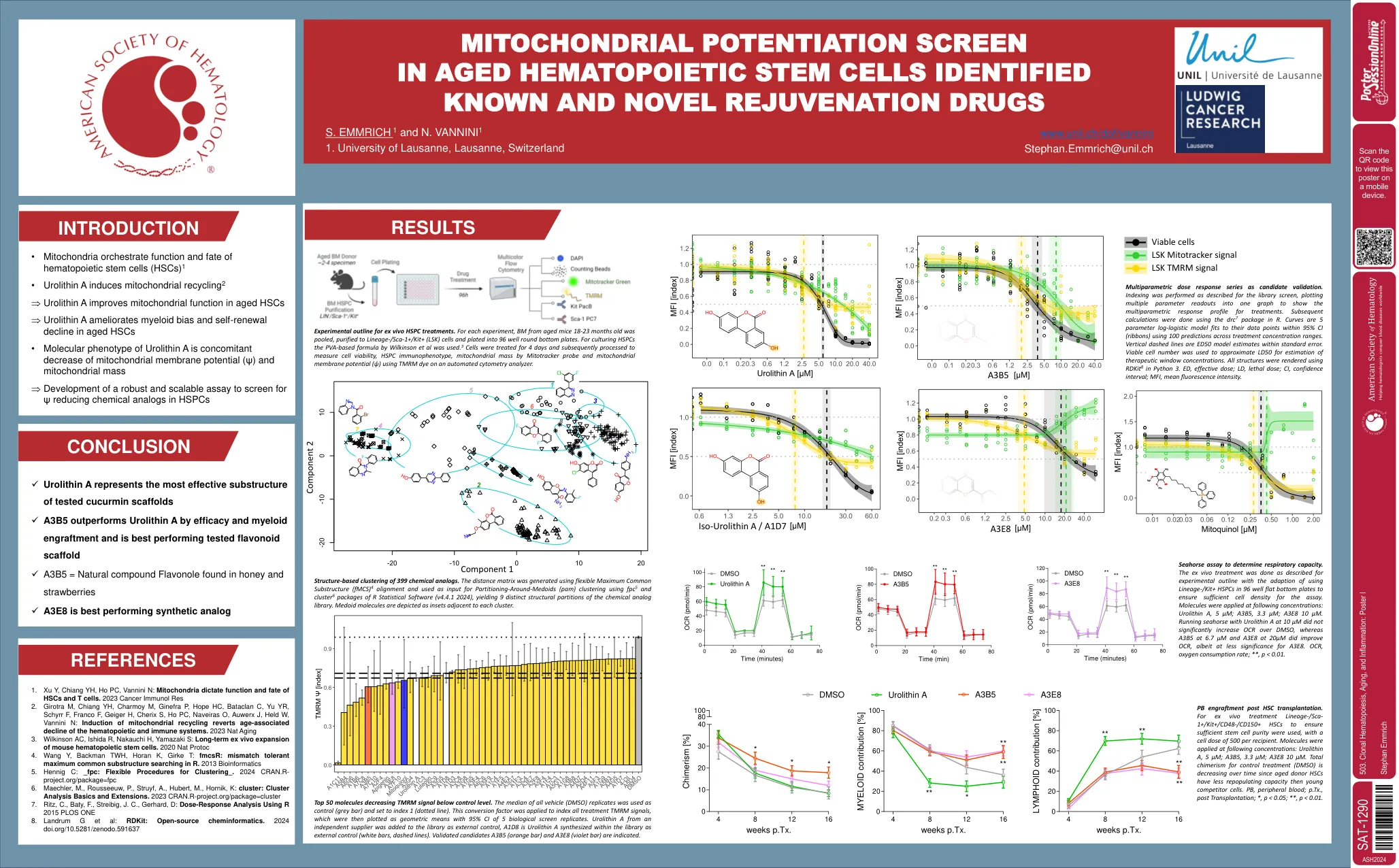
503。克隆造血,衰老和炎症:海报I
1。Xu Y,Chiang YH,HO PC,Vannini N:线粒体决定HSC和T细胞的功能和命运。2023 CANCAR IMMUNOL RES 2。Girotra M, Chiang YH, Charmoy M, Ginefra P, Hope HC, Bataclan C, Yu YR, Schyrr F, Franco F, Geiger H, Cherix S, Ho PC, Naveiras O, Auwerx J, Held W, Vannini N: Induction of mitochondrial recycling reverts age-associated decline of the hematopoietic and immune系统。2023 NAT老化3。Wilkinson AC,Ishida R,Nakauchi H,Yamazaki S:小鼠造血干细胞的长期离体扩张。 2020 NAT ProtoC 4。 Wang Y,Backman TWH,Horan K,Girke T:FMCSR:不匹配的最大最大常见子结构搜索R. 2013 Bioinformatics 5。 Hennig C:_FPC:clustering_的灵活过程。 2024 cran.r- project.org/package=fpc 6。 Maechler,M.,Rousseeuw,P.,Struyf,A.,Hubert,M.,Hornik,K:集群:聚类分析基础知识和扩展。 2023 cran.r-project.org/package=cluster 7。 Ritz,C.,Baty,F.,Streibig,J.C.,Gerhard,D:使用R 2015 PLOS ONE 8。的剂量反应分析 Landrum G等人:RDKIT:开源化学信息学。 2024 doi.org/10.5281/zenodo.591637Wilkinson AC,Ishida R,Nakauchi H,Yamazaki S:小鼠造血干细胞的长期离体扩张。2020 NAT ProtoC 4。Wang Y,Backman TWH,Horan K,Girke T:FMCSR:不匹配的最大最大常见子结构搜索R. 2013 Bioinformatics 5。Hennig C:_FPC:clustering_的灵活过程。2024 cran.r- project.org/package=fpc 6。Maechler,M.,Rousseeuw,P.,Struyf,A.,Hubert,M.,Hornik,K:集群:聚类分析基础知识和扩展。2023 cran.r-project.org/package=cluster 7。Ritz,C.,Baty,F.,Streibig,J.C.,Gerhard,D:使用R 2015 PLOS ONE 8。Landrum G等人:RDKIT:开源化学信息学。2024 doi.org/10.5281/zenodo.591637

关于克隆造血(CH)
您的医生将进行检查以检查血液癌。这些测试可能包括血液检测,例如全血细胞计数(CBC)或基因检测。血液计数低,遗传突变的某些类型和大小会增加患血癌的风险。基因检测有助于监测突变的类型和大小。大多数CH患者患血癌的风险非常低。

造血细胞在治疗中的造血细胞移植
从广义上讲,有两种类型的造血细胞移植(HCT,以前在本政策中称为造血干细胞移植[HSCT]),自体和同种异体。自体HCT的目的是治疗疾病(例如淋巴瘤)伴有骨髓剂量的化学疗法(有或没有放射线),它们具有反对该疾病的活性。接受者自己的HCT(以前收集)在化学疗法后注入,以重新建立正常的骨髓功能。在同种异体移植中,受体在骨髓疗法或非毛囊治疗后从供体中接收HCT,以重新建立正常的骨髓功能,并将新的血液系统用作免疫疗法的平台,这就是一种所谓的“移植物与肿瘤”的效果。造血细胞可以从骨髓,外周血或脐带血液中收获,不久后新生儿分娩后不久。尽管脐带血是一种同种异体来源,但其中的细胞在抗原上是“幼稚的”,因此与排斥反应或移植物抗宿主病(GVHD)的发生率较低有关。
