XiaoMi-AI文件搜索系统
World File Search System酸化

使用 TiOx 作为电阻层的电阻存储器的制造
图 3 ReRAM 特性的电极依赖性:(a) 50×50 μm 2 ,(b) 200×200 μm 2 。 5.结论我们利用 TiO x 作为电阻变化层制作了 ReRAM,并评估了其特性。在本次创建的条件下,没有观察到复位操作。这被认为是因为在复位操作过程中,由于氧气的释放,灯丝没有断裂。比较电极尺寸,50×50 μm2 的较小元件与 200×200 μm2 的元件相比,可获得更优异的特性。这被认为表明了氧化退火过程中的尺寸依赖性。 6.参考文献 [1] A. Hardtdegen 等,IEEE Transactions on Electron Devices,第 65 卷,第 8 期,第 3229-3236 页 (2018) [2] Takeo Ninomiya,基于氧化物材料设计和可靠性建模的电阻式存储器量产,名古屋大学研究生院博士论文 (2016) [3] D.Carta 等,ACS Appl. Mater. Interfaces,第 19605-19611 页 (2016) [4] D. Acharyya 等,微电子可靠性。54,第 541-560 页 (2014)。

气候与国防战略 - 武装部队部
气候变化导致环境缓慢而渐进的变化,例如变暖和海平面上升、海洋酸化和缺氧、由于降水模式变化而导致某些地区干旱、或传染病的出现和传播。它也是自然灾害突然发生的原因,正如热浪、洪涝灾害、海洋淹没现象和气旋等极端天气事件趋于增多和加剧所表明的那样。据世界气象组织统计,过去五十年里,它们的数量增加了五倍。
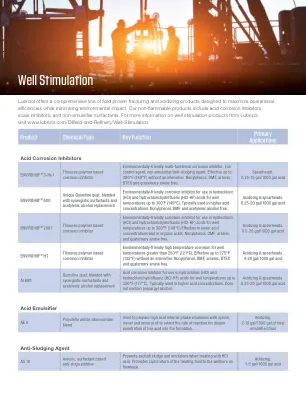

HACCP 计划示例
• HACCP 团队和检查员需要了解产品是什么、如何制造(成分 + 加工步骤)、使用哪种容器以及如何运输和消费。 • 成分可能决定产品的类型(酸性/酸化;低 Aw 等) • 保质期长与易腐烂 • 玻璃容器与塑料容器 • 单份、RTE/RTD 或分份

母校StudiorumUniversitàdiBologna Archivio ...
Biagi,E.,Caroselli,E.,Barone,M.,Pezzimenti,M.,Teixido,N.,Soverini,M。等。(2020)。微生物组组成中的模式与居住在二氧化碳通风口的地中海珊瑚珊瑚钙化钙化的钙化钙化室中的海洋酸化不同。总环境科学,724,1-11 [10.1016/j.scitotenv.2020.138048]。

气候防御战略 - 武装部队部
气候变化导致环境缓慢而渐进地改变,例如气候变暖和海平面上升、海洋酸化和缺氧、由于降水状况变化而导致某些地区干旱化或传染病的出现和传播。这也是自然界突然出现暴力行为的原因,正如热浪、洪涝灾害以及海洋沉没和气旋等极端天气事件呈增多和加剧趋势所表明的那样。据世界气象组织统计,过去五十年里,它们的数量增加了五倍。

TAQ DNA聚合酶
TAQ DNA聚合酶是一种重组94 kDa DNA聚合酶,该聚合酶在带有克隆的Thermu Aquaticus DNA聚合酶基因的大肠杆菌菌株中表达。它具有5'-3'聚合酶和外切核酸酶活性,并且没有可检测到的3'-5'外切核酸酶活性。TAQ DNA聚合酶的延伸速率为1-2 kb/ min。此外,它具有3'腺苷酸化活性。因此,PCR产品可直接用于TA-CLONing程序。

衰老神经生物学
唾液酸结合免疫球蛋白样凝集素 (Siglec) 受体与神经退行性过程有关,但唾液酸在生理性衰老中的作用仍未完全了解。我们研究了唾液酸生物合成所必需的葡萄糖胺-2-表异构酶/N-乙酰甘露糖胺激酶 (GNE + / ) 杂合子小鼠脑内唾液酸化降低的影响。我们证明 GNE + / 小鼠在 6 个月时不同脑区已出现唾液酸化降低、海马体突触减少、小胶质细胞树突状化减少,随后 12 个月时神经元丢失增加。转录组分析显示未发现促炎症变化,这表明在衰老过程中 GNE + / 小鼠的突触和神经元被固有的稳态免疫过程所清除。与补体 C3 缺乏的小鼠杂交挽救了早期发生的神经元和突触丢失以及小胶质细胞树突状化的变化。因此,糖萼的唾液酸有助于大脑稳态,并充当大脑先天免疫系统的识别系统。2020 作者。由 Elsevier Inc. 出版。这是一篇根据 CC BY-NC-ND 许可协议开放获取的文章(http://creativecommons.org/licenses/by-nc-nd/4.0/)。

鱼免疫能力足以面对气候变化?
持续的气候变化已经与野生鱼类和养殖鱼类的疾病爆发增加有关。在这里,我们评估了当前关于气候变化相关的生态免疫学的知识,重点是探索多种压力源的交互作用,重点是临时,缺氧,盐度和酸化。我们的文献综述表明,温度和溶解氧的急性和慢性变化会损害鱼类免疫力,从而导致疾病易感性增加。此外,已经证明温度和缺氧可以增强某些病原体/寄生虫的感染并加速疾病进展。也很少有针对酸化的研究,但是直接的免疫作用似乎受到限制,而盐度研究导致了对比结果。同样,对于揭示同时改变环境因素的相互作用所必需的多压力实验仍然很少。这最终阻碍了我们估计气候变化在多大程度上会妨碍鱼类免疫力的能力。我们对表观遗传调节机制的评论突出了鱼类免疫反应对不断变化的环境的适应潜力。但是,由于表观遗传学研究数量有限,因此无法得出总体结论。最后,我们提供了如何更好地估计鱼类未来免疫研究的现实气候变化情景影响的前景。

节能研究部
随着能源的加速,需要开发高效的能量转换技术。我们的小组专注于高温运行能量转化设备,并集中在固体氧化物燃料电池(SOFC)上,这些燃料电池(SOFC)以高效的方式将各种燃料(例如化石燃料和生物量)转化为电能,以及固体氧化物电解电池(SOEC),这些电池(SOEC),这些电池(SOEC),这些电池(SOEC)将造成的能量添加到较高的能量中,添加了高效果,添加了高级效果,高效率。特别是,在SOFC提高耐用性和可靠性的努力中,我们在与公司和大学合作中作为公共研究机构发挥了重要作用。将来,我们将通过对更高效的能源转换设备进行全面的研究和发展,包括开发用于固态离子学现象的评估技术,从而为低碳社会实现。