XiaoMi-AI文件搜索系统
World File Search System髓鞘
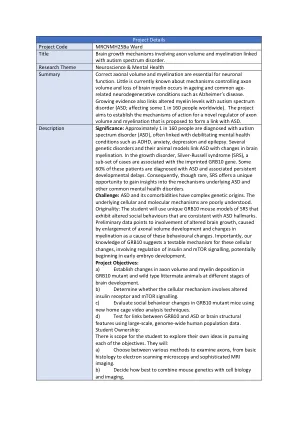
项目详情项目代码 MRCNMH25Ba 病房标题涉及轴突体积和髓鞘形成的大脑生长机制与自闭症谱系障碍相关。
项目详情 项目代码 MRCNMH25Ba 病房标题 涉及轴突体积和髓鞘形成的大脑生长机制与自闭症谱系障碍有关。 研究主题 神经科学与心理健康 摘要 正确的轴突体积和髓鞘形成对神经元功能至关重要。目前对控制轴突体积的机制知之甚少,大脑髓鞘的流失发生在衰老和常见的与年龄相关的神经退行性疾病,如阿尔茨海默病中。越来越多的证据还将髓鞘水平改变与自闭症谱系障碍(ASD;影响全球每 160 人中约 1 人)联系起来。该项目旨在确定一种新型轴突体积和髓鞘形成调节剂的作用机制,该调节剂被认为与 ASD 有联系。描述意义:大约每 160 人中就有 1 人被诊断患有自闭症谱系障碍 (ASD),通常与多动症、焦虑、抑郁和癫痫等使人衰弱的精神健康疾病有关。几种遗传性疾病及其动物模型将 ASD 与大脑髓鞘形成的变化联系起来。在生长障碍 Silver-Russell 综合征 (SRS) 中,一部分病例与印记的 GRB10 基因有关。这些患者中约有 60% 被诊断患有 ASD 并伴有持续发育迟缓。因此,尽管罕见,SRS 提供了一个独特的机会来深入了解 ASD 和其他常见精神健康障碍的潜在机制。挑战:ASD 及其合并症具有复杂的遗传起源。潜在的细胞和分子机制尚不清楚。原创性:学生将使用独特的 GRB10 SRS 小鼠模型,这些模型表现出与 ASD 特征一致的改变的社会行为。初步数据表明,由于轴突体积发育增大和髓鞘形成变化导致大脑生长改变,这是导致这些行为变化的原因。重要的是,我们对 GRB10 的了解表明这些细胞变化存在可测试的机制,涉及调节胰岛素和 mTOR 信号传导,可能始于早期胚胎发育。项目目标:a) 确定 GRB10 突变体和野生型同窝动物在大脑发育的不同阶段的轴突体积和髓鞘沉积变化。b) 确定细胞机制是否涉及改变的胰岛素受体和 mTOR 信号传导。c) 使用新的家笼视频分析技术评估 GRB10 突变小鼠的社会行为变化。d) 使用大规模全基因组人类群体数据测试 GRB10 与 ASD 或大脑结构特征之间的联系。学生所有权:学生在追求每个目标时都有机会探索自己的想法。他们将:a) 选择各种方法来检查轴突,从基本的组织学到电子扫描显微镜和复杂的 MRI 成像。b) 决定如何最好地将小鼠遗传学与细胞生物学和成像相结合,


失调的胆碱能信号传导抑制脱髓鞘后的少突胶质细胞
少突胶质细胞祖细胞(OPC)募集和少突胶质细胞分化的失调导致人类脱髓鞘疾病(如多发性硬化症(MS))中的再髓呈失败。毒蕈碱受体的缺失增强了OPC分化和再生。然而,配体依赖性信号传导与本构受体激活的作用尚不清楚。我们假设脱髓鞘后失调的乙酰胆碱(ACH)释放有助于配体介导的激活阻碍髓磷脂修复。在慢性丘陵(CPZ)诱导的脱髓鞘(雄性和雌性小鼠)之后,我们观察到ACH浓度增加了2.5倍。ACH浓度的这种增加可以归因于ACH合成或乙酰胆碱酯酶 - /丁酰胆碱酯酶(BCHE)介导的降解降低。使用胆碱乙酰基转移酶(CHAT)记者小鼠,我们识别出在Lysolecithin和CPZ脱髓鞘后增加了CHAT-GFP的表达。CHAT-GFP表达在载脂内溶血素诱导的脱髓鞘后的受伤和未受伤的轴突的子集中上调。在CPZ-甲状腺call体中,在GFAP +星形胶质细胞和轴突中观察到CHAT-GFP,这表明神经元和星形胶质细胞ACH释放的潜力。CPZ脱髓鞘后,cpz call体的BCHE表达显着降低。这种减少是由于骨髓少突胶质细胞的丧失,这是BCHE的主要来源。我们确定成熟的少突胶质细胞密度的剂量依赖性降低,对OPC募集没有影响。确定溶血石注射后配体介导的毒蕈碱信号传导的作用,我们给予了胆碱酯酶抑制剂Neostigine,以人为提高ACH。一起,这些结果支持了脱髓鞘后配体介导的毒蕈碱受体激活的功能作用,并表明ACH稳态失调直接导致MS中的再生性失败。

第2阶段的疗效和安全性,riliprubart,一种C1抑制剂,在慢性炎症性脱髓鞘性多神经病中
1个神经肌肉疾病部门,神经病学系,西班牙巴塞罗那市De la Santa Creu I Sant Pau医院; Centro Para larespjitionaciónBioMédicaEnRed Enfermedades Raras(Ciberer),西班牙马德里; 2 Cedars Sinai Medical Center,美国加利福尼亚州洛杉矶; 3德国杜塞尔多夫海因里希海大学医学院神经病学系;悉尼分校,悉尼,澳大利亚,悉尼分校的大脑和思维中心;奥地利维也纳医科大学神经病学系;捷克共和国Olomouc Palacky University Olomouc神经病学系; 4荷兰鹿特丹大学医学中心Erasmus MC; 5赛诺菲研发,美国马萨诸塞州剑桥市神经病学发展; 6美国赛诺菲; 7赛诺菲研发,美国新泽西州布里奇沃特的生物统计学和编程; 8英国伦敦伦敦大学学院UCL皇后广场神经病学研究所

大麻二醇减少脑室内出血脑损伤,保留髓鞘并防止血液屏障功能障碍
脑室内出血(IVH)是极早产儿的长期残疾的重要原因,没有目前的治疗。这项研究评估了使用未成熟大鼠在IVH模型中大麻二醇(CBD)的潜在神经保护作用。ivh通过左室周围注射裂术胶原酶在1天大的(P1)Wistar大鼠中诱导。一些大鼠会产前接受CBD(10 mg/kg I.P.到达大坝),然后是5 mg/kg I.P.IVH后 6、30和54 h(IVHÞCBD,n¼30)。 其他IVH大鼠接收了车辆(IVH车,n¼34)和用车辆处理的非IVH大鼠用作对照(SHM,n¼29)。 大鼠在P6,P14或P45处被人性地杀死。 Brain damage (motor and memory performance, area of damage, Lactate/N-acetylaspartate ratio), white matter injury (ipsi- lateral hemisphere and corpus callosum volume, oligodendroglial cell density and myelin basic protein signal), blood-brain barrier (BBB) integrity (Mfsd2a, occludin and MMP9 expression, gadolinium leakage), in fl然后评估了弹性毒性(TLR4,NFκB和TNFα表达,在促炎细胞的纤维化中),兴奋毒性(谷氨酸/N-乙酰基 - 吐型比)和氧化应激(蛋白质硝基化)。 cbd阻止了IVH的长期运动和认知后果,在短期和长期保护的寡头细胞中减少了脑损伤,从而保留了足够的髓鞘形成并保持BBB的完整性。 CBD的保护作用与炎症,兴奋性和氧化应激的调节有关。 CBD是改善IVH诱导的未成熟脑损伤的潜在候选者。6、30和54 h(IVHÞCBD,n¼30)。其他IVH大鼠接收了车辆(IVH车,n¼34)和用车辆处理的非IVH大鼠用作对照(SHM,n¼29)。大鼠在P6,P14或P45处被人性地杀死。Brain damage (motor and memory performance, area of damage, Lactate/N-acetylaspartate ratio), white matter injury (ipsi- lateral hemisphere and corpus callosum volume, oligodendroglial cell density and myelin basic protein signal), blood-brain barrier (BBB) integrity (Mfsd2a, occludin and MMP9 expression, gadolinium leakage), in fl然后评估了弹性毒性(TLR4,NFκB和TNFα表达,在促炎细胞的纤维化中),兴奋毒性(谷氨酸/N-乙酰基 - 吐型比)和氧化应激(蛋白质硝基化)。cbd阻止了IVH的长期运动和认知后果,在短期和长期保护的寡头细胞中减少了脑损伤,从而保留了足够的髓鞘形成并保持BBB的完整性。CBD的保护作用与炎症,兴奋性和氧化应激的调节有关。CBD是改善IVH诱导的未成熟脑损伤的潜在候选者。总而言之,在未成熟的大鼠中,CBD降低了IVH诱导的脑损伤及其短期和长期后果,显示出强大的和多效性神经保护作用。

髓鞘在神经退行性疾病中的作用:对药物靶点和神经保护策略的影响
摘要:中枢神经系统轴突的髓鞘化具有许多优点,包括减少信号传输的能量消耗和提高信号速度。轴突周围的髓鞘由少突胶质细胞形成的多层膜组成,而特定的糖蛋白和脂质在这一形成过程中发挥着各种作用。髓鞘虽然有益,但其失调和退化可能会造成损害。炎症、氧化应激以及细胞代谢和细胞外基质的变化可导致这些轴突脱髓鞘。这些因素是某些脱髓鞘疾病(包括多发性硬化症)的标志性特征。脱髓鞘的影响也与青光眼和阿尔茨海默病等疾病的原发性退化以及继发性退化过程有关。这揭示了髓鞘与神经退行性变的次级过程之间的关系,包括创伤性损伤和跨突触变性后的退化。髓鞘在原发性和继发性退化中的作用也是探索髓鞘再生策略和目标的兴趣所在,包括使用抗炎分子或纳米颗粒来输送药物。尽管在髓鞘再生动物模型中使用这些方法

渗透性脱髓鞘是低钠血症校正的并发症:系统评价
背景:低钠血症的快速纠正,尤其是在严重和慢性的情况下,可能导致渗透性脱粒化。诊断和治疗低钠血症的最后一个指南(2014年)建议校正限制为10 meq/l/天。我们的目标是总结已发表的渗透性脱髓鞘案例,以评估该建议的充分性。方法:渗透性脱髓鞘病例报告的系统审查。,我们以1997年至2019年在1997年至2019年发表的英语或葡萄牙语中出版的18岁以上的图像或病理解剖学案例包括确认的病例。结果:我们评估了96例渗透脱髓鞘,女性的58.3%,平均年龄为48.2±12.9岁。中位入院钠为105 MEQ/L,> 90%的患者患有严重的低钠血症(<120 MEQ/L)。胃肠道疾病(38.5%),埃菲主义(31.3%)和利尿剂使用(27%)的报告很常见。低钠血症矫正主要用等渗盐水(46.9%)或高渗(33.7%)进行。相关的低钙矫正发生在18.8%。在66.6%的病例中,住院第一天对纳特血症的纠正高于10 meq/l;速度没有报道为22.9%,只有10.4%的速度小于10 meq/ l/天。结论:在50岁以下的患有严重低钠血症和快速纠正的女性中,渗透性脱髓鞘化的发展主要是主导。在10.4%的病例中,即使校正<10 meq/l/day也存在脱髓化。这些数据加强了对高风险患者的保守目标的需求,例如4-6 meq/l/天,不超过8 meq/l/day的极限。

髓鞘形成和轴突直径的发育,实现快速而精确的动作电位传导
图 2 蒙古沙鼠梯形体 (TB) 髓鞘的高分辨率图像。抗神经丝相关抗原 (3A10) (a – c) 和神经丝重链 (NFH) (d – f) 的抗体用作轴突标记物。抗髓鞘碱性蛋白 (MBP) 的抗体显示髓鞘。在出生后第 6 天 (a、a')、出生后第 9 天 (b、b 0 ) 和出生后第 13 天 (c、c 0 ) 从 TB 区域沙鼠大脑冠状振动切片中获取共聚焦单光学图像。在出生后第 7 天 (d、d 0 )、出生后第 10 天 (e、e') 和出生后第 14 天 (f、f 0 ) 从 TB 区域沙鼠大脑矢状振动切片中获取共聚焦单光学切片。 (a – f) 轴突标记物 3A10 (a – c) 和 NFH (d – f) 以红色显示,MBP 免疫反应性以绿色表示。(a' – f 0) 相应的 MBP 染色图像。出生后第 6 天 (a、a') 可以看到短的、有时是点状的髓鞘碎片,其间散布着较长的无髓鞘间隙。少突胶质细胞 (白色星号) 积极产生 MBP,用抗 MBP 抗体标记。在出生后第 7 天的矢状切面中,可以看到 TB 纤维的横截面。一小部分 TB 轴突被 MBP 包围,用抗 MBP 抗体标记。出生后第 9 天 (b、b 0),TB 中的大部分轴突都是髓鞘化的。然而,人们可以很容易地注意到一些轴突没有被髓鞘包裹 (白色箭头,b)。在出生后第 10 天的矢状切面上,大多数轴突被髓鞘包裹,但有些没有(白色箭头,e)。到出生后第 13 天(c,c 0)可以看到髓鞘包裹所有轴突。请注意,髓鞘轴突排列非常紧密,以至于很难勾勒出属于单个轴突的髓鞘。在出生后第 14 天的矢状切面上,TB 区域的所有轴突横截面都被髓鞘包裹。比例尺:20 μ m。

对因 TUBB4A 突变导致基底神经节和小脑萎缩、髓鞘形成不足的青少年进行深部脑刺激:示例 c
在出生后的第一年,他的运动和发育里程碑正常,但随后在 18 个月大时出现肌张力低下和平衡问题。他在 5 岁时出现右下肢肌张力障碍,并开始逐渐丧失粗大和精细运动功能,包括独立行走的能力。他在 7 岁时发展为全身性肌张力障碍。考虑手术时的表型表现主要是肌张力过高,四肢活动范围有限,无法负重,脊柱侧弯严重。他的肌张力障碍对肠内苯海索、巴氯芬、卡比多巴-左旋多巴、亚叶酸钙和丁苯那嗪也有抵抗力。根据他父母的报告,在植入一个

微生物在脱髓鞘疾病的疗法发生中的作用
摘要:多发性硬化症(MS),神经霉素炎(NMO)和髓磷脂少突胶质细胞糖蛋白抗体疾病(MOGAD)是中枢神经系统(CNS)的炎症性疾病,具有多因素病因。环境因素对其发展很重要,微生物可能发挥决定性作用。他们可以直接损坏CNS,但是它们与免疫系统的相互作用更为重要。涉及的可能机制包括分子模仿,表位扩散,旁观者激活和双细胞受体理论。已经明确确定了爱泼斯坦 - 巴尔病毒(EBV)在MS中的作用,因为血清阳性是MS发作的必要条件。 EBV与遗传和环境因素相互作用,例如低水平的维生素D和人内源性逆转录病毒(HERV),这是另一种涉及该疾病的微生物。在感染结核分枝杆菌,EBV和人类免疫障碍病毒后,已经描述了许多神经瘤谱障碍(NMOSD)的发作或加剧病例。但是,尚未发现与病毒确定的关系。对幽门螺杆菌的螺旋细菌提出了可能的作用,尤其是在水通道蛋白4抗体的个体中。感染后可能发生摩根摩加人的发作,主要是在疾病的单相过程中。已经假设了HERV在Mogad中的角色。在这篇综述中,我们研究了对传染因素在MS,NMO和Mogad中的参与的当前理解。我们的目标是阐明每种微生物在引发疾病并影响其临床进展中的作用。我们的目的是讨论具有良好作用的感染因素,以及在各种研究中产生相互影响结果的传染性因素。