XiaoMi-AI文件搜索系统
World File Search SystemAAV8
正在进行的国际阶段I/II ...
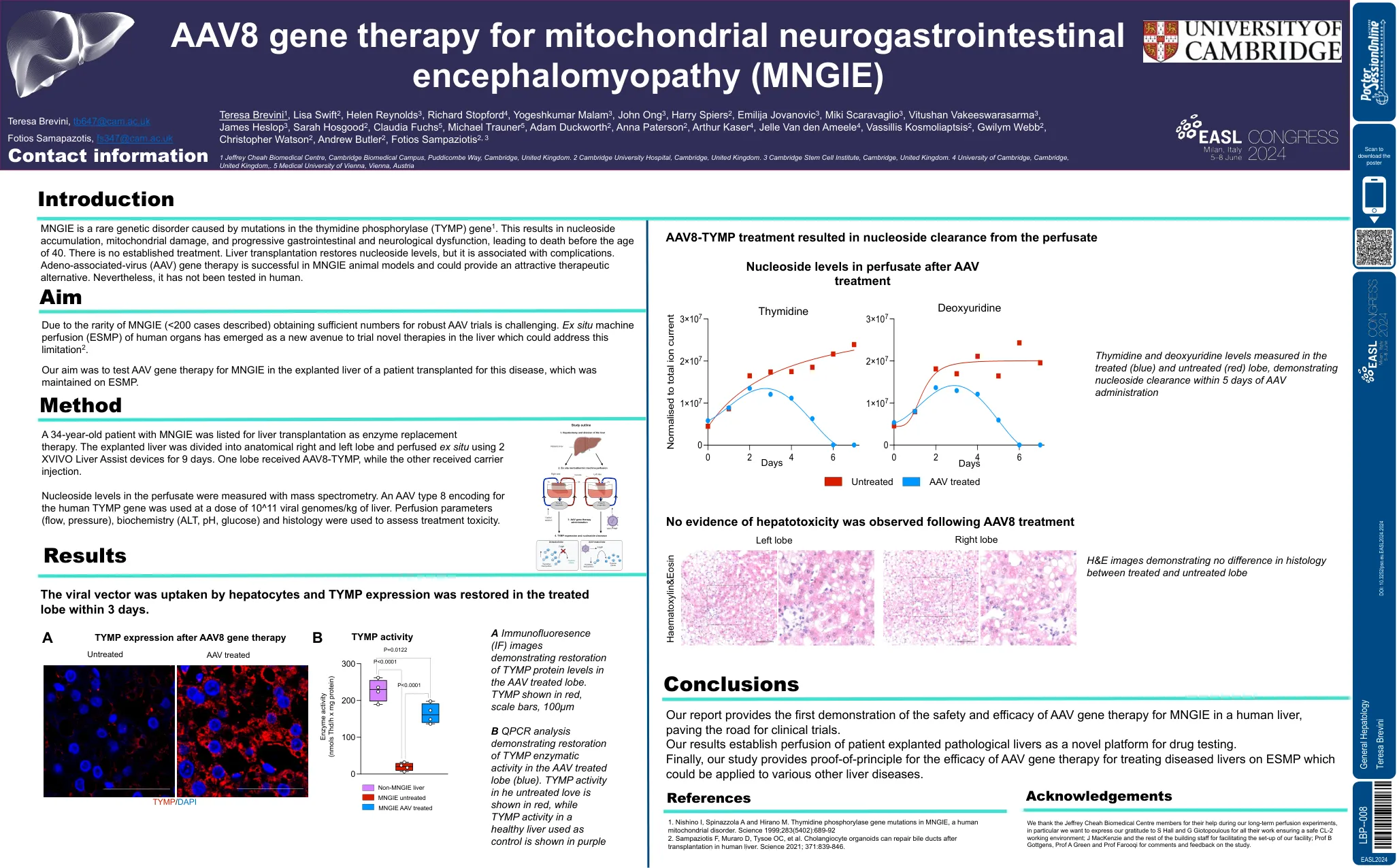
mngie是一种罕见的遗传疾病,是由胸苷磷酸化酶(TYMP)基因1中的突变引起的。这会导致核苷的积累,线粒体损伤以及进行性胃肠道和神经功能障碍,导致40岁之前死亡。没有既定的治疗方法。肝移植恢复核苷水平,但与并发症有关。腺相关病毒(AAV)基因疗法在MNGIE动物模型中成功,并且可以提供有吸引力的治疗方法。尽管如此,它尚未在人类中进行测试。

电脉冲暴露减少了转导HEPG2细胞所需的AAV8剂量
我们证明,在存在增强的绿色荧光蛋白(EGFP)表达腺体相关病毒(AAV8)载体的情况下,将电场脉冲在体外施加到肝细胞,使给定的传输水平与HISPATOCYCYP相比,将腺相关的病毒量(AAV8)降低了50-折叠式的电气量不超过50-折叠量。 接触。我们在标准井板中的8个暴露条件下进行了48个实验观测。电脉冲暴露涉及具有375 V/ cm场强度的单个80-MS脉冲。我们的研究表明,电脉冲暴露会导致细胞中EGFP的表达增强,这表明转导效率提高。如果成功地转换为体内环境,在我们的研究中观察到的增强转导将是一种有希望的迹象,证明了AAV载体所需剂量的潜在减少。对电场脉冲对体外AAV转导的影响是重要的前一步。

AAV8 衍生的 CRISPR 对乙肝病毒的抑制... - NET
慢性乙型肝炎病毒 (HBV) 感染的治愈性治疗仍是一个遥远的目标,HBV 复制过程中稳定的共价闭合环状 DNA (cccDNA) 的持续存在是目前批准用于治疗 HBV 的药物难以突破的关键障碍。由于基因组编辑的准确性、效率和成本效益,CRISPR/Cas 技术被广泛应用于基因治疗和抗病毒策略。虽然 CRISPR/Cas 可能清除 cccDNA,但确保其安全性是应用的必要条件。在我们的研究中,我们分析了几种启动子的肝脏特异性,并构建了 CRISPR/金黄色葡萄球菌 Cas9 (SaCas9) 系统结合肝嗜性 AAV8(其中 AAV 指腺相关病毒)的候选启动子来验证对抗 HBV 的功效。结果显示,将原始启动子替换为肝脏特异性启动子的重建 CRISPR/SaCas9 系统在体内和体外仍然可以抑制 HBV 复制。 3种功能性向导RNA(gRNA)T 2 、T 3 和T 6 针对不同HBV基因型的保守区域,在不同肝脏特异性启动子的作用下均表现出较好的抗HBV效果,且3种gRNA对A、B、C基因型HBV的复制均有不同程度的抑制作用。在EnhII-Pa1AT启动子和AAV8作用下,SaCas9在其他器官或组织中的表达较肝脏进一步降低。本研究结果有助于确保CRISPR/Cas9系统的作用局限于肝脏,从而降低因非特异性靶向其他器官而产生不良有害作用的可能性,为肝脏的临床应用提供参考。

AAV8基因治疗线粒体神经胃肠道...702。CAR-T细胞疗法侵略性B细胞淋巴瘤
•基于关键Zuma-1试验4,Axibabtagene Ciololeucel(AXI-CEL)是第一个针对LBCL的成年患者批准的嵌合抗原受体(CAR)T-CELL疗法,这些患者是复发/难治性(R/R)至2+疗法(众多)(众多)(大量)(众多)(美国)和欧洲(US)和欧洲(US)和欧洲(US)和欧洲(US)和欧洲(US)。5,6在Zuma-7试验7成功之后,美国食品和药物管理局和欧洲药品局将这种迹象扩展到2L使用。

对HeartQol健康相关的生活质量问卷的验证提出了14个项目问卷的验证概述,旨在成为我们
AAV2I8,腺相关病毒AAV2/AAV8载体capsid; CMV,巨细胞病毒; DNA,脱氧核糖核酸; I-1C,组成性活性抑制剂1; EMA,欧洲药品局; FDA,美国食品和药物管理局。1。Pathak A等。Circ Res。 2005; 96(7):756-766。 2。 Asokan A等。 nat生物技术。 2010; 28:79-82。 3。 Ishikawa K等。 Circ Res。 2018; 123:601-613。Circ Res。2005; 96(7):756-766。 2。 Asokan A等。 nat生物技术。 2010; 28:79-82。 3。 Ishikawa K等。 Circ Res。 2018; 123:601-613。2005; 96(7):756-766。2。Asokan A等。nat生物技术。2010; 28:79-82。 3。 Ishikawa K等。 Circ Res。 2018; 123:601-613。2010; 28:79-82。3。Ishikawa K等。 Circ Res。 2018; 123:601-613。Ishikawa K等。Circ Res。 2018; 123:601-613。Circ Res。2018; 123:601-613。2018; 123:601-613。

分泌的PD-L1通过局部和全身AAV基因治疗减轻小鼠的炎症性关节炎
结果:在表达SPD-L1(SHPD-L1)最佳亚型的AAV6载体的关节内注射后,与野生型PD-L1相比,观察到更大的效力,而关节炎改善所需的AAV6/SHPD-L1则较低。为了研究SPD-L1的全身表达的治疗作用,我们通过恢复轨道注射在CIA小鼠中施用AAV8/SHPD-L1基因治疗,并发现关节炎症和PAW肿胀的显着改善,在幼稚的小鼠中表现出相似的表型。在AAV8/ SHPD-L1处理的CIA小鼠中,总免疫球蛋白和抗胶原蛋白的特定抗体的水平低于对照组的抗体。在SHPD-L1处理的小鼠中,血液中促炎性细胞因子的水平也显着降低。此外,脾脏中的T细胞凋亡率在处理后的小鼠中增加了2倍。最后,我们通过肌肉注射研究了AAV/ SHPD-L1的治疗作用。注射AAV6/SHPD-L1后,pAW肿胀减少,关节炎症减少,血液中促炎细胞因子的降低和降低。SHPD-L1的治疗作用是通过用AAV载体肌肉处理来依赖于剂量的。

腺相关病毒载体的表征
60- 61 图 1.6:AAV8 X 预测的失活基因同源物。62 图 1.7:重组 AAV 包装系统。66 图 1.8:AAV REP 结合位点。83 图 3.1:纯化 AAV 中污染物序列的扩增。105 图 3.2:AAV 污染物扩增子的 DNASE 抗性。106 图 3.3:污染物研究中使用的重组 AAV 包装系统的 REP 结合位点位置。

GMO 注册号 G0876
请参阅 GMO 申请,GMO 注册号 G0876-01,用于研究 ITL-3001-CL-101,该申请已于 2024 年 12 月 19 日获得机构批准。Intellia Therapeutics, Inc.(Intellia,赞助商/GMO 通知人)特此通知机构,由于优先考虑其他赞助商项目,撤回临床试验 ITL-3001-CL-101,用于研究产品 NTLA-3001。NTLA-3001 是一种基于 AAV8 和 CRISPR/Cas9 的体内基因编辑研究疗法,用于治疗患有 alpha-1 抗胰蛋白酶缺乏症 (AATD) 相关肺病的成年人。

BAd-CRISPR:在成年小鼠肩胛间棕色脂肪组织中诱导基因敲除
CRISPR/Cas9 已实现多种组织中的可诱导基因敲除;然而,尚未有其在棕色脂肪组织 (BAT) 中的应用报道。在此,我们开发了棕色脂肪细胞 CRISPR (BAd-CRISPR) 方法来快速检测一个或多个基因的功能。使用 BAd-CRISPR,将表达单向导 RNA (sgRNA) 的腺相关病毒 (AAV8) 直接施用于在棕色脂肪细胞中表达 Cas9 的小鼠的 BAT。我们表明,将 AAV8-sgRNA 局部施用于成年小鼠的肩胛间 BAT 可强有力地转导棕色脂肪细胞,并使脂联素、脂肪甘油三酯脂肪酶、脂肪酸合酶、周脂素 1 或硬脂酰辅酶 A 去饱和酶 1 的表达降低 90% 以上。施用多个 AAV8 sgRNA 可同时敲除多达三个基因。 BAd-CRISPR 诱导移码突变并抑制靶基因 mRNA 表达,但不会导致 BAT 中脱靶突变的大量积累。我们利用 BAd-CRISPR 创建了可诱导的解偶联蛋白 1 (Ucp1) 敲除小鼠,以评估 UCP1 缺失对成年小鼠适应性产热的影响。可诱导的 Ucp1 敲除不会改变核心体温;然而,BAd-CRISPR Ucp1 小鼠的成纤维细胞生长因子 21 循环浓度升高,并且 BAT 基因表达发生变化,与通过增加过氧化物酶体脂质氧化而产生的热量一致。其他分子适应性预示着额外的细胞效率低下,蛋白质合成和周转增加,线粒体对线粒体编码基因表达的依赖降低,核编码线粒体基因表达增加。这些数据表明 BAd-CRISPR 是一种加速脂肪组织生物学发现的有效工具。

AAV8-hUGT1A1 与雷帕霉素联合治疗新生儿感染的疗效...
一项使用腺相关病毒血清型 8 (AAV8)-人尿苷二磷酸葡萄糖醛酸转移酶 1A1 (h UGT1A1) 治疗遗传性严重未结合高胆红素血症 (Crigler-Najjar 综合征) 的临床试验正在进行中,但临床前数据表明,由于在生长中的肝脏中肝细胞增殖时转基因表达丧失,该疗法在儿童中的长期疗效不佳。本研究旨在确定在相关动物模型中在什么年龄可以获得长期疗效,以及免疫调节是否允许使用相同的 AAV 载体进行重复治疗。新生、哺乳期和幼年 Ugt1a1 缺乏大鼠接受了临床相关剂量的 AAV8-h UGT1A1 ,并监测血清胆红素水平和血清中的抗 AAV8 中和抗体 (NAbs)。使用雷帕霉素方案研究了预防对载体的免疫反应的可能性,该方案从载体给药前 2 天开始,到给药后 21 天结束,每天进行腹膜内 (ip) 注射。在出生后第 1 天 (P1) 或 P14 接受治疗的大鼠中,12 周后校正 (部分) 消失,而在 P28 注射的大鼠中校正保持稳定。将初始载体给药与免疫抑制方案相结合可防止雌性大鼠诱导 NAbs,从而至少可以部分有效地进行再次给药。再次注射后无法防止 NAbs 的诱导,这表明该策略对于先前存在的抗 AAV NAbs 水平较低的患者无效。