XiaoMi-AI文件搜索系统
World File Search SystemBUD

ashs_vol-113_box-01 _Book-05
放热炎症米卡利亚河中的冷损伤。Amer。 J. Enol。 葡萄。 31:1-6。 Proebsting,E.L.,Jr.1963。 空气温度和芽的作用在确定休眠的“ Elberta”桃果芽的坚固性中。 proc。 Amer。 Soc。 hort。 SCI。 83:259-269。 Proebsting,E.L.,Jr.1970。 秋天和冬季温度与落叶果树的花芽行为和木质硬度的关系。 Hortscience 5:422-424。 proebsting,E.L。和H.H. 工厂。 1972。 比较了“ Bing”樱桃和“ Elberta”桃子的果芽中坚硬的反应。 J. Amer。 Soc。 hort。 SCI。 97:802-806。 Quamme,H.A。 1978。 在越冬的桃花芽中过冷的机制。 J. Amer。 Soc。 hort。 SCI。 103:57-61。 Quamme,H.A。 1983。 空气温度与水含量的关系和越冬的桃花芽的过冷。 J. Amer。 Soc。 hort。 SCI。 108:697-701。 Quamme,H.A。 1985。 通过深冷的避免木质植物中的冷冻损伤。 Acta Hort。 168:11-30。 Quamme,H.A.,C。Stushnoff和C.J. Weiser。 1972。 关系Amer。J. Enol。 葡萄。 31:1-6。 Proebsting,E.L.,Jr.1963。 空气温度和芽的作用在确定休眠的“ Elberta”桃果芽的坚固性中。 proc。 Amer。 Soc。 hort。 SCI。 83:259-269。 Proebsting,E.L.,Jr.1970。 秋天和冬季温度与落叶果树的花芽行为和木质硬度的关系。 Hortscience 5:422-424。 proebsting,E.L。和H.H. 工厂。 1972。 比较了“ Bing”樱桃和“ Elberta”桃子的果芽中坚硬的反应。 J. Amer。 Soc。 hort。 SCI。 97:802-806。 Quamme,H.A。 1978。 在越冬的桃花芽中过冷的机制。 J. Amer。 Soc。 hort。 SCI。 103:57-61。 Quamme,H.A。 1983。 空气温度与水含量的关系和越冬的桃花芽的过冷。 J. Amer。 Soc。 hort。 SCI。 108:697-701。 Quamme,H.A。 1985。 通过深冷的避免木质植物中的冷冻损伤。 Acta Hort。 168:11-30。 Quamme,H.A.,C。Stushnoff和C.J. Weiser。 1972。 关系J. Enol。葡萄。31:1-6。Proebsting,E.L.,Jr.1963。空气温度和芽的作用在确定休眠的“ Elberta”桃果芽的坚固性中。proc。Amer。 Soc。 hort。 SCI。 83:259-269。 Proebsting,E.L.,Jr.1970。 秋天和冬季温度与落叶果树的花芽行为和木质硬度的关系。 Hortscience 5:422-424。 proebsting,E.L。和H.H. 工厂。 1972。 比较了“ Bing”樱桃和“ Elberta”桃子的果芽中坚硬的反应。 J. Amer。 Soc。 hort。 SCI。 97:802-806。 Quamme,H.A。 1978。 在越冬的桃花芽中过冷的机制。 J. Amer。 Soc。 hort。 SCI。 103:57-61。 Quamme,H.A。 1983。 空气温度与水含量的关系和越冬的桃花芽的过冷。 J. Amer。 Soc。 hort。 SCI。 108:697-701。 Quamme,H.A。 1985。 通过深冷的避免木质植物中的冷冻损伤。 Acta Hort。 168:11-30。 Quamme,H.A.,C。Stushnoff和C.J. Weiser。 1972。 关系Amer。Soc。hort。SCI。 83:259-269。 Proebsting,E.L.,Jr.1970。 秋天和冬季温度与落叶果树的花芽行为和木质硬度的关系。 Hortscience 5:422-424。 proebsting,E.L。和H.H. 工厂。 1972。 比较了“ Bing”樱桃和“ Elberta”桃子的果芽中坚硬的反应。 J. Amer。 Soc。 hort。 SCI。 97:802-806。 Quamme,H.A。 1978。 在越冬的桃花芽中过冷的机制。 J. Amer。 Soc。 hort。 SCI。 103:57-61。 Quamme,H.A。 1983。 空气温度与水含量的关系和越冬的桃花芽的过冷。 J. Amer。 Soc。 hort。 SCI。 108:697-701。 Quamme,H.A。 1985。 通过深冷的避免木质植物中的冷冻损伤。 Acta Hort。 168:11-30。 Quamme,H.A.,C。Stushnoff和C.J. Weiser。 1972。 关系SCI。83:259-269。 Proebsting,E.L.,Jr.1970。 秋天和冬季温度与落叶果树的花芽行为和木质硬度的关系。 Hortscience 5:422-424。 proebsting,E.L。和H.H. 工厂。 1972。 比较了“ Bing”樱桃和“ Elberta”桃子的果芽中坚硬的反应。 J. Amer。 Soc。 hort。 SCI。 97:802-806。 Quamme,H.A。 1978。 在越冬的桃花芽中过冷的机制。 J. Amer。 Soc。 hort。 SCI。 103:57-61。 Quamme,H.A。 1983。 空气温度与水含量的关系和越冬的桃花芽的过冷。 J. Amer。 Soc。 hort。 SCI。 108:697-701。 Quamme,H.A。 1985。 通过深冷的避免木质植物中的冷冻损伤。 Acta Hort。 168:11-30。 Quamme,H.A.,C。Stushnoff和C.J. Weiser。 1972。 关系83:259-269。Proebsting,E.L.,Jr.1970。秋天和冬季温度与落叶果树的花芽行为和木质硬度的关系。Hortscience 5:422-424。proebsting,E.L。和H.H.工厂。1972。比较了“ Bing”樱桃和“ Elberta”桃子的果芽中坚硬的反应。J. Amer。 Soc。 hort。 SCI。 97:802-806。 Quamme,H.A。 1978。 在越冬的桃花芽中过冷的机制。 J. Amer。 Soc。 hort。 SCI。 103:57-61。 Quamme,H.A。 1983。 空气温度与水含量的关系和越冬的桃花芽的过冷。 J. Amer。 Soc。 hort。 SCI。 108:697-701。 Quamme,H.A。 1985。 通过深冷的避免木质植物中的冷冻损伤。 Acta Hort。 168:11-30。 Quamme,H.A.,C。Stushnoff和C.J. Weiser。 1972。 关系J. Amer。Soc。hort。SCI。 97:802-806。 Quamme,H.A。 1978。 在越冬的桃花芽中过冷的机制。 J. Amer。 Soc。 hort。 SCI。 103:57-61。 Quamme,H.A。 1983。 空气温度与水含量的关系和越冬的桃花芽的过冷。 J. Amer。 Soc。 hort。 SCI。 108:697-701。 Quamme,H.A。 1985。 通过深冷的避免木质植物中的冷冻损伤。 Acta Hort。 168:11-30。 Quamme,H.A.,C。Stushnoff和C.J. Weiser。 1972。 关系SCI。97:802-806。Quamme,H.A。1978。在越冬的桃花芽中过冷的机制。J. Amer。 Soc。 hort。 SCI。 103:57-61。 Quamme,H.A。 1983。 空气温度与水含量的关系和越冬的桃花芽的过冷。 J. Amer。 Soc。 hort。 SCI。 108:697-701。 Quamme,H.A。 1985。 通过深冷的避免木质植物中的冷冻损伤。 Acta Hort。 168:11-30。 Quamme,H.A.,C。Stushnoff和C.J. Weiser。 1972。 关系J. Amer。Soc。hort。SCI。 103:57-61。 Quamme,H.A。 1983。 空气温度与水含量的关系和越冬的桃花芽的过冷。 J. Amer。 Soc。 hort。 SCI。 108:697-701。 Quamme,H.A。 1985。 通过深冷的避免木质植物中的冷冻损伤。 Acta Hort。 168:11-30。 Quamme,H.A.,C。Stushnoff和C.J. Weiser。 1972。 关系SCI。103:57-61。Quamme,H.A。1983。空气温度与水含量的关系和越冬的桃花芽的过冷。J. Amer。 Soc。 hort。 SCI。 108:697-701。 Quamme,H.A。 1985。 通过深冷的避免木质植物中的冷冻损伤。 Acta Hort。 168:11-30。 Quamme,H.A.,C。Stushnoff和C.J. Weiser。 1972。 关系J. Amer。Soc。hort。SCI。 108:697-701。 Quamme,H.A。 1985。 通过深冷的避免木质植物中的冷冻损伤。 Acta Hort。 168:11-30。 Quamme,H.A.,C。Stushnoff和C.J. Weiser。 1972。 关系SCI。108:697-701。Quamme,H.A。1985。通过深冷的避免木质植物中的冷冻损伤。Acta Hort。168:11-30。 Quamme,H.A.,C。Stushnoff和C.J. Weiser。 1972。 关系168:11-30。Quamme,H.A.,C。Stushnoff和C.J.Weiser。1972。关系

仅敲除 LincRNA#1 不会影响凯尔特 POLLED 等位基因杂合牛的无角表型
长基因间非编码 RNA(lincRNA#1)在无角牛胎儿的角芽区过度表达,表明其可能在角芽抑制中发挥作用。使用基因组编辑来测试此序列的缺失是否与角表型有关。将两个具有高突变效率的 gRNA 靶向 lincRNA#1 序列两侧的 5′ 和 3′ 区域,在授精后 6 小时与 Cas9 一起作为核糖核蛋白复合物注射到牛受精卵(n = 121)中。在产生的囊胚(n = 31)中,84% 具有预期的 3.7 kb 缺失;在这些具有 3.7 kb 缺失的胚胎中,88% 是双等位基因敲除。将 39 个推测已编辑的 7 天囊胚移植到 13 头同步受体母牛体内,导致 10 例妊娠,其中 5 个胚胎在 POLLED 基因座上为显性 PC POLLED 等位基因杂合子,5 个胚胎为隐性 pp 基因型。产生的胎儿中有 8 个 (80%) 为双等位基因 lincRNA#1 敲除,其余两个为嵌合体。RT-qPCR 分析用于确认敲除胎儿中不存在 lincRNA#1 表达。对基因型 (PC p) POLLED 、lincRNA#1 敲除胎儿的表型和组织学分析显示,其形态与未编辑的对照无角胎儿相似,表明仅缺乏 lincRNA#1 不会导致有角表型。


代谢组谱图揭示了不同茶品种质量的组成差异
抽象茶厂在生物活性化合物中丰富,包括类黄酮,氨基酸,生物碱,萜类化合物和脂质,这些主要影响茶质量和口味。尽管有许多关于不同茶品种的代谢产物的研究,但其生物合成和调节的组成差异仍然是未知的。在这项研究中,使用靶向的代谢组学广泛的代谢组学,包括192个黄酮和28 neminds和28 amino,从根尖的芽中检测到505种代谢产物('shuchazao':'scz':'scz':'scz':'huangkui':'hk'和'hk'和'zijuan':'zj':'zj'。代谢产物分析表明,黄酮醇和花色苷主要以三种品种的糖苷形式分布,其中花青素及其糖苷主要在“ ZJ”中积累,表明与颜色属性有相关性。EGCG成为三种品种中最丰富的Flavan-3-ols化合物。l-茶氨酸代表主要的游离氨基酸,与1叶相比,主要集中在顶端芽中,但同样,脂质与游离氨基酸相似,主要是在三个品种的顶端芽中积聚。这些发现为遗传和代谢物多样性提供了宝贵的见解,从而增强了我们对茶叶特定代谢物的生物合成的理解。

final-cts_very-mild and-mild-assthma-cpg.pdf
摘要背景:此哮喘指南更新的重点是在频谱的轻度末端对哮喘患者的管理。它适用于1岁及以上和成人的儿童。此更新旨在解决该患者组中的新临床试验,以及全球哮喘(GINA)哮喘战略组的建议的变化。本指南将当前的证据应用于加拿大的情况。方法:代表性的多学科专家小组进行了正式的临床实践指南开发过程。根据患者/问题,干预,比较,结果(PICO)方法,总共定义了9个关键临床问题。小组进行了基于证据的系统文献综述,评估并对相关证据进行了分级,以综合11个关键建议。这些建议是在存在的加拿大哮喘指南的背景下进行了审查的,并强调了先前准则的变化。结果:更新的证据表明,与非常轻度和轻度哮喘的人相比,与PRN SABA相比,每天吸入的皮质类固醇(ICS)þPRN短期β-激动剂(SABA)减少了加重并改善哮喘控制。在12岁的儿童中有新的证据,与PRN SABA相比,PRN BUDESONIDE/FORMOTEROL(BUD/FORM)的成年人降低了加重,在患有轻度哮喘的患者中,有不同水平的证据。在患有轻度哮喘的个体中,仍然建议将每日ICSþPRNSABA作为第一线控制器疗法。患有非常轻度的哮喘患者应为较高的加重风险,可以选择从prn saba转换为每日iCsþprnsaba(所有年龄段)或prn芽/形式(12岁)。然而,在12岁的个体中,尽管大量哮喘教育和支持,但对每日药物的依从性较差,PRN芽/形式是另一种选择。间歇性地使用非常高剂量的IC来急性丧失哮喘控制的急性损失,而学龄前儿童则可能会受到伤害。讨论:本指南提供了对证据的详细审查,并为在加拿大背景下为学龄前儿童,儿童和成人提供了非常轻度和温和的哮喘的建议。加拿大胸腔协会2021年哮喘指南更新将与以前的指南合并这些建议,以提供解决哮喘诊断和管理的文件。
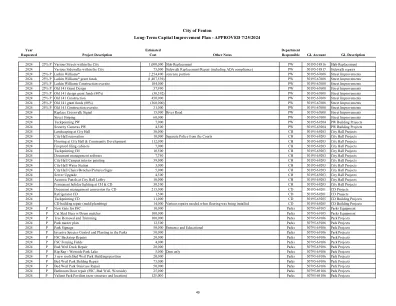
芬顿市长期基础设施改善计划
2024 25% P 城市内各条街道 1,000,000 板更换 PW 50195-58816 板更换 2024 城市内各条人行道 75,000 人行道更换/维修(包括 ADA 合规性) PW 50195-58817 人行道维修 2024 25% P Larkin Williams* 2,254,400 混凝土部分 PW 50195-67608 街道改善 2024 25% P Larkin Williams* 补助资金(1,407,539) PW 50195-67608 街道改善 2024 25% P Larkin Williams 施工现场监督 104,000 PW 50195-67608 街道改善 2024 25% P 旧 141 补助设计37,990 PW 50195-67608 街道改善 2024 25% P 旧 141 设计补助资金 (80%) (30,392) PW 50195-67608 街道改善 2024 25% P 旧 141 建设 450,000 PW 50195-67608 街道改善 2024 25% P 旧 141 补助资金 (80%) (360,000) PW 50195-67608 街道改善 2024 25% P 旧 141 建设现场 31,000 PW 50195-67608 街道改善 2024 更换人行横道信号 15,000 河路 PW 50195-67608 街道改善 2024 街道标线60,000 PW 50195-67608 街道改善 2024 勾缝 PW 7,000 PW 50195-69204 PW 建筑项目 2024 安全摄像头 PW 4,500 PW 50195-69204 PW 建筑项目 2024 市政厅景观美化 30,000 CH 50195-69203 市政厅项目 2024 市政厅翻新 10,000 将警察与法院分开 CH 50195-69203 市政厅项目 2024 市政厅地板和社区发展 112,000 CH 50195-69203 市政厅项目 2024 防火文件柜 7,000 CH 50195-69203 市政厅项目 2024 勾缝 CH 10,500 CH 50195-69203 市政厅项目 2024 文档管理软件 7,750 CH 50195-69203 市政厅项目 2024 市政厅校园内部绘画 39,000 CH 50195-69203 市政厅项目 2024 市政厅供水站 3,000 CH 50195-69203 市政厅项目 2024 市政厅椅子/长凳/图片/标志 5,000 CH 50195-69203 市政厅项目 2024 服务器升级 4,500 CH 50195-69203 市政厅项目 2024 市政厅大厅隔音板 10,000 CH 50195-69203 市政厅项目 2024 CH 和 CD 的永久节日灯饰30,500 CH 50195-69203 市政厅项目 2024 文件管理转换 CD 215,000 CD 50195-69201 CD 项目 2024 冰箱 CD 1,500 CD 50195-69201 CD 项目 2024 填缝 CD 11,000 CD 50195-69205 CD 建筑项目 2024 CD 建筑维修(模具/管道) 14,000 安装地板时需要进行的各种维修 CD 50195-69205 CD 建筑项目 2024 P FSC 的新大门 10,000 公园 50795-69105 公园设备 2024 P 带滚筒粉碎机的 Cat 滑移转向装载机 100,000 公园 50795-69105 公园设备2024 P 树木移除和修剪 100,000 公园 50795-69106 公园项目 2024 P 公园总体规划 12,500 公园 50795-69106 公园项目 2024 P 公园标牌 50,000 入口和教育公园 50795-69106 公园项目 2024 P 公园入侵物种控制和种植 50,000 公园 50795-69106 公园项目 2024 P FSC 挡土墙修复 20,000 公园 50795-69106 公园项目 2024 P FSC 击剑场 4,000 公园 50795-69106 公园项目 2024 P Bud Weil 码头修复 20,000 公园 50795-69106 公园项目 2024 P 护堤 - 西侧公园湖 5,000 仅水坝 公园 50795-69106 公园项目 2024 P 3 个新屋顶 Bud Weil 公园建筑/展馆 20,000 公园 50795-69106 公园项目 2024 P Bud Weil 公园建筑修复 75,000 公园 50795-69106 公园项目 2024 P Bud Weil 公园楼梯修复 50,000 公园50795-69106 公园项目 2024 P 浴室门维修 (FSC, Bud Weil, Westside) 25,000 公园 50795-69106 公园项目 2024 P Valiant Park Pavilion (新结构和位置) 125,000 公园 50795-69106 公园项目


伯氏锥虫的线粒体 RNA 编辑
2 启示 3 4 5 Evgeny S. Gerasimov a# 、 Dmitry A. Afonin a# 、 Oksana A. Korzhavina a 、 Julius Lukeš b,c 、 Ross Low d 、 Neil Hall d 、 6 Kevin Tyler e 、 Vyacheslav Yurchenko f * 、 Sara L. Zimmer g * 7 8 9 a 分子生物学系,罗蒙诺索夫莫斯科国立大学,莫斯科 119234,俄罗斯 10 jalgard@gmail.com (ESG)、afoninmsu@outlook.com (DAA)、korzhavina.oksana.bio.msu@gmail.com 11 (OAK) 12 13 b 寄生虫学研究所,生物中心,捷克科学院,370 05 捷克布杰约维采,捷克14 jula@paru.cas.cz (JL) 15 16 c 南波西米亚大学科学学院,370 05 捷克布杰约维采,捷克 17 18 d 厄尔汉姆研究所,诺维奇研究园区,诺维奇 NR4 7UZ,英国 19 Ross.Low@earlham.ac.uk(RL),neil.hall@earlham.ac.uk(NH) 20 21 e 诺维奇医学院,东英吉利亚大学,诺维奇 NR4 7TJ,英国 22 K.Tyler@uea.ac.uk(KT) 23 24 f 生命科学研究中心,俄斯特拉发大学科学学院,710 00 俄斯特拉发,捷克 25 vyacheslav.yurchenko@osu.cz(VY) 26 27 g 明尼苏达大学医学院,德卢斯校区,明尼苏达州德卢斯,55812,美国 28 szimmer3@d.umn.edu (SLZ) 29 30 # 平等贡献 31 * 通信地址 32 33

2018 年 TUKE 活动报告
科希策技术大学 (TUKE) 为周边地区提供科技知识库、创新和劳动力,以塑造有益且可持续的未来和提高公民的生活质量。科希策科技大学通过大学各院系所有科学领域的创新研究和卓越教育实现了这一目标。

花朵表面附生细菌的生长模式
花朵中寄生着各种附生细菌群落,这些细菌会影响花朵的功能、传粉媒介相互作用以及植物的整体适应性。然而,人们对这些细菌的丰度如何随着花朵的衰老而变化以及这些变化与花朵寿命的关系知之甚少。在本研究中,我研究了从开花期(花蕾开放到花朵)到衰老期(花朵枯萎)的花朵生命周期中细菌丰度的变化,并探索了对花朵寿命的潜在影响。我们通过确定两个野外季节中 8 种植物花朵的平均衰老年龄来追踪花朵的年龄。花蕾在开花前被标记,使我们能够从花蕾开放的时刻(标志着花朵开花的开始)到可见枯萎的开始(表明衰老的开始)追踪花朵的寿命,我们通过平板计数确定了花朵表面可培养细菌的丰度,并测量了环境温度、湿度和降水如何影响这些模式。我们的结果表明,随着花朵的衰老,它们会积累细菌,寿命较短的花朵通常比寿命较长的花朵积累细菌的速度更快。然而,与预期相反,附生细菌的丰度与花朵寿命无关,这表明附生细菌可能不会直接影响花朵寿命。相反,环境条件起着重要作用;温度升高与细菌丰度降低有关,而湿度升高则支持细菌丰度增加和花朵寿命延长。这些发现表明,花朵上的细菌丰度可能受外部因素影响,而对花朵寿命没有直接影响,这凸显了花朵衰老与环境条件之间复杂的相互作用。