XiaoMi-AI文件搜索系统
World File Search SystemCRRNA

基于设计的,双组分指南RNA
合成生物学的目标之一是实现具有可编程输入和输出的任意分子回路的设计。这样的电路桥接了电子和自然电路的性质,以可预测的活细胞内处理信息。基因组编辑是合成分子回路的潜在强大组成部分,无论是用于调节靶基因的表达还是稳定地记录信息到基因组DNA。然而,编程分子事件,例如蛋白质 - 蛋白质相互作用或作为基因组编辑触发因素诱发的接近性仍然具有挑战性。在这里,我们演示了一种称为P3编辑的策略,该策略将蛋白质 - 蛋白质接近性与功能性CRISPR-CAS9双组分指南RNA的形成联系起来。通过工程化crRNA:tracrrna相互作用,我们证明了各种已知的蛋白质 - 蛋白质相互作用以及化学诱导的蛋白质结构域的二聚化,可用于激活人类细胞中的素数编辑或基础编辑。此外,我们探讨了P3编辑如何结合基于ADAR的RNA传感器的输出,有可能允许特定的RNA在较大电路中诱导特定的基因组编辑。我们的策略增强了基于CRISPR的基因组编辑的可控性,从而促进了其在活细胞中部署的合成分子电路中的使用。

CRISPR-Cas13a系统的应用进展
成簇的规律间隔短回文重复序列 (CRISPR) 和 CRISPR 相关 (Cas) 蛋白是一种适应性免疫系统,可保护原核生物和一些感染原核生物的病毒免受外来核酸(如病毒和质粒)的侵害。大多数古细菌和大约一半的细菌的基因组都含有各种 CRISPR-Cas 系统。CRISPR-Cas 系统依赖于 CRISPR RNA (crRNA)。它们充当导航系统,通过识别入侵的外来核酸并结合 Cas 蛋白来特异性地切割和破坏外来核酸。在本综述中,我们简要概述了 CRISPR-Cas 系统的进化和分类,重点介绍了 CRISPR-Cas13a 系统的功能和应用。我们描述了 CRISPR-Cas13a 系统并讨论了其 RNA 指导的核糖核酸酶功能。同时,我们简要介绍了CRISPR-Cas13a系统的作用机制,并总结了CRISPR-Cas13a系统在病原体检测、真核生物、农业、生物传感器和人类基因治疗中的应用。我们对CRISPR-Cas13a的正确理解已经得到拓宽,CRISPR-Cas13a系统将有助于开发新的RNA靶向工具。因此,了解CRISPR-Cas13a效应蛋白的结构、功能和生物学特性的基本细节对于优化RNA靶向工具至关重要。

文章抑制 SHROOM1 可通过促进同源定向修复来改善体内和体外基因整合
摘要:同源重组 (HR) 常用于实现靶向基因整合,因为与微同源介导的末端连接 (MMEJ) 或非同源末端连接 (NHEJ) 相比,它具有更高的精度和可操作性。由于它只在细胞分裂期间发生,因此似乎对动物细胞和胚胎中的基因整合效率较低。在这里,我们开发了全基因组高通量筛选和随后的配对 crRNA 文库筛选,以寻找抑制同源定向修复 (HDR) 的基因。我们发现,在报告系统中,敲低 SHROOM1 的 HDR 细胞比对照细胞富集多达 4.7 倍。无论供体类型如何,下调 SHROOM1 均可显著促进人类和小鼠细胞中成簇的规律间隔短回文重复序列 (CRISPR)/CRISPR 相关蛋白 9 核酸酶 (Cas9) 切割后的基因整合。通过在微注射过程中应用 SHROOM1 siRNA,小鼠胚胎的敲入效率也可以加倍。HEK293T 细胞中 SHROOM1 缺失引起的 HDR 效率增加可被 HDR 抑制剂 YU238259 抵消,但不能被 NHEJ 抑制剂抵消。这些结果表明 SHROOM1 是一个 HDR 抑制基因,SHROOM1 敲低策略可能适用于多种应用,包括基因编辑以生成用于研究基因功能和人类疾病的细胞系和动物模型。

鉴定元素组对EH CRISPR -CAS9系统及其在基因组工程中的应用
抽象的非编码RNA(CRRNA)是由定期间隔短的短膜重复序列(CRISPR)基因座(CRISPR)基因座和与CRISPR相关的(CAS)的(CAS)的蛋白形成的,形成了crispr-CAS系统形成的复合物,这些复合物形成了与CAS-CataLe creeavage crre creage creeavage crrecry crrecry creeaveage crecrecry crrecry creeavage物质相互作用的相互作用的相互作用的creve criss cospectes。这些核糖蛋白启用的核酸易于编程的靶向促进了基于CRISPR的分子生物学工具的实施,用于体内和DNA和RNA靶标的体外修饰。尽管到目前为止鉴定出靶向DNA的Cas核酸酶的多样性,但在Pyogenes链球菌SPCAS9的天然和工程衍生物是基因组工程中最广泛使用的,至少部分是由于它们的CATA-LECALLYTIC稳健性以及一个异常短的图案(5''-ngg-ngg-3'''Pam)farnk sequence。但是,SPCAS9变体的大尺寸会损害该工具向真核细胞的传递,并且需要较小的替代品。在这里,我们在元基因组中识别与较小的CAS9蛋白(EHCAS9)相关的新的CRISPR-CAS9系统,该系统靶向以5'-NGG-3'PAM为两侧的DNA序列。我们开发了一种简化的EHCAS9工具,该工具专门切割DNA靶标,可用于原核生物和真核细胞中的基因组编辑应用。

CRISPR-Cas12a 辅助 Amycolatopsis mediterranei 基因组编辑
Amycolatopsis mediterranei U32 是利福霉素 SV 的工业生产者,其衍生物长期以来一直是抗分枝杆菌的一线药物。为了对这种重要的工业菌株进行基因改造,过去几十年来人们付出了很多努力,我们实验室成功开发了一种基于同源重组的方法,但该方法需要使用抗生素抗性基因进行正向选择,并且不支持方便的无标记基因删除。在本研究中,我们利用成簇的规律间隔短回文重复序列 (CRISPR) 系统在 A. mediterranei U32 中建立了基因组编辑系统。具体来说,土拉弗朗西斯菌亚种。 novicida Cas12a (FnCas12a) 基因首先整合到 U32 基因组中,在 CRISPR RNA (crRNA) 的指导下产生靶向特异性双链 DNA (dsDNA) 断裂 (DSB)。然后,DSB 可以通过非同源 DNA 末端连接 (NHEJ) 系统或同源定向修复 (HDR) 途径修复,分别在靶基因中产生不准确或准确的突变。除了 A. mediterranei 之外,本研究还可能为 Amycolatopsis 属其他物种中 CRISPR 辅助基因组编辑系统的开发提供启示。

ArciTect™ Cas9-eGFP 核酸酶
ArciTect™ Cas9-eGFP 核酸酶是一种融合蛋白,由增强型绿色荧光蛋白 (eGFP) 和来自化脓性链球菌的野生型 Cas9 重组蛋白组成;它包含一个 C 端连接的 eGFP 分子。ArciTect™ Cas9-eGFP 核酸酶需要与向导 RNA(例如 ArciTect™ sgRNA(目录号 #200-0013)或由 ArciTect™ tracrRNA(目录号 #76016)和 ArciTect™ crRNA(目录号 #76010)组成的双链)结合,以形成核糖核蛋白 (RNP) 复合物。该 RNP 复合物在基因组中的位点特定位置产生双链断裂。 ArciTect™ Cas9-eGFP 核酸酶还在 N 端含有核定位信号,确保 RNP 复合物转位至细胞核,从而提高基因组编辑的效率。由于 RNP 复合物在转染后完全发挥作用,因此在转位至细胞核后可立即发挥作用。RNP 复合物在 48 小时内降解,为基因组编辑提供了充足的时间,同时减少了 RNP 复合物持续存在可能导致的脱靶效应。使用 RNP 系统还可以避免生成稳定的 Cas9 表达细胞系的繁琐过程,从而节省时间并降低由于可诱导表达系统泄漏而导致脱靶效应的风险。化脓性链球菌 Cas9 使用原间隔区相邻基序 (PAM) 序列 NGG(其中 N 可以是任何核苷酸)。如果靶序列下游没有基因组 PAM 位点,酶就不会裂解。

细菌CRISPR/CAS9系统作为所有健康问题的有希望的解决方案,并进步的生物工程
缩写:BP1,肿瘤抑制剂p53结合蛋白1; BRCA,乳腺癌抗原;汽车,嵌合抗原受体; CAS9,CRISPR相关蛋白9;级联,抗病毒防御的CRISPR综合体; CMR,CAS模块坡道(重复相关的神秘蛋白质); CMR III-B,多个亚基III型B CRISPR RNA-CAS蛋白; CPF1,Prevotella和Francisella1的CRISPR; CRISPR,定期间隔间隔室; Crrna,Crispr RNA; CSM III-A,多支亚基III-A CRISPR-CAS蛋白; dcas9/ sgrna-sg I,停用cas9/短指南RNA-Sybrr-green i; DNA-PK,DNA-蛋白K; DNA-PKC,DNA蛋白K催化亚基; DSB,双链断裂; ege,额外的基因元素; GRNA,导向RNA; HDR,同源性维修; IAP,碱性磷酸酶同工酶; MRE 11,减数分裂重组11; NHEJ,非同理结局加入; PAM,原始间隔者相邻基序; PD,程序性细胞死亡; RAD,重组酶A;代表,重复的外部回文; RPA,复制蛋白A; RT,逆转录酶; Sgrna,简短的指南RNA; SSB,单链断裂; tracrrna,反式激活CRISPR RNA; XLF,类似XRCC4的因子; XRCC 4,X射线修复交叉补充蛋白4; Yoyo-1,(恶唑黄色)
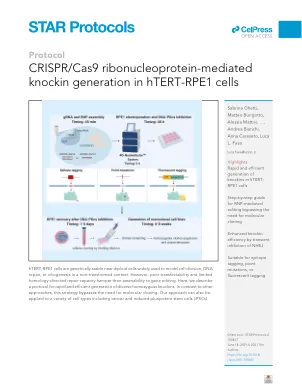
CRISPR/CAS9核糖核蛋白介导的HTERT-RPE1细胞中的敲除蛋白
图1。使用ssDNA或PCR产物作为HDR模板(a)上部的蛋白质标记的策略示例:据报道编码Centriolar远端附属物蛋白SCLT1的C-末端的基因组序列。带下划线的序列代表CrRNA识别位点,PAM序列为黄色,垂直虚线表示切割位点。鉴于距离内源性终止密码子(BOLD大写字母的TAA)距离为14 bp,插入位点被任意定位为距剪切位点1 bp的位置,即在SCLT1的密码子之间的最接近的交界处。在下部,密码子(上图)和相应的氨基酸性残基(下图)构成了插入物:蓝色大写字母是指可易加的链接器,然后是v5-tag(红色)和附加的外源终止密码子(黑色)。50 bp lha或rha = 50碱基对左同源臂或右同源臂。(b)使用PCR产物作为供体DNA生成具有荧光蛋白(FP)的蛋白质(您最喜欢的蛋白质,YFP)的C端标记的示意图。PCR模板由带有FP,2A元件和电阻盒(R)的标准质粒(左侧)组成。使用一对60mer引物进行PCR反应。在右侧代表了目标基因座(您最喜欢的基因,YFG)的编辑。

利用多种 V 型 CRISPR 进行无 PAM 诊断
V 型 CRISPR-Cas 效应子通过促进核酸生物标志物的检测,彻底改变了分子诊断。然而,它们依赖于目标双链 DNA (dsDNA) 上原间隔区相邻基序 (PAM) 位点的存在,这极大地限制了它们作为诊断工具的灵活性。在这里,我们提出了一种名为 PICNIC 的新方法,该方法只需对当代 CRISPR 检测方案进行约 10 分钟的简单修改,即可解决基于 CRISPR 的诊断的 PAM 问题。我们的方法包括通过高温和高 pH 处理将 dsDNA 分离成单个单链 DNA (ssDNA) 链。然后,我们以无 PAM 的方式使用多种 Cas12 酶检测释放的 ssDNA 链。我们通过成功将 PICNIC 用于 Cas12 家族的三种不同亚型(Cas12a、Cas12b 和 Cas12i)的无 PAM 检测,展示了 PICNIC 的实用性。值得注意的是,通过将 PICNIC 与含有 crRNA 的截短 15 核苷酸间隔区相结合,我们证明了使用 CRISPR 进行临床上重要的单核苷酸多态性的 PAM 独立检测。我们采用这种方法检测 HIV-1 的耐药变体(特别是 K103N 突变体)的存在,该变体在突变附近缺乏 PAM 位点。此外,我们成功地将我们的方法应用于临床样本,通过在 HCV 基因组中无 PAM 位点以 100% 特异性检测和基因分型 HCV-1a 和 HCV-1b 变体。总之,PICNIC 是一种简单但具有突破性的方法,它通过消除 PAM 序列的限制提高了基于 CRISPR-Cas12 的诊断的灵活性和精确度。

利用 CRISPR-Cas9 基因编辑弗里德赖希共济失调患者的造血干细胞
弗里德赖希共济失调 (FRDA) 是一种常染色体隐性神经退行性疾病,由 frataxin (FXN) 基因内含子 1 中的 GAA 重复扩增引起,导致线粒体铁结合蛋白 frataxin 的表达显著降低。我们之前报告说,同基因造血干细胞和祖细胞 (HSPC) 移植可防止 FRDA 小鼠模型 YG8R 中的神经退行性。我们表明,挽救机制是由功能性 frataxin 从 HSPC 衍生的小胶质细胞/巨噬细胞转移到神经元/肌细胞所介导的。在本研究中,我们报告了使用 CRISPR-Cas9 系统进行 FRDA 自体 HSPC 移植的第一步。我们首次鉴定出一对 CRISPR RNA(crRNA),它们可有效消除人类 FRDA 淋巴母细胞中的 GAA 扩增,恢复 frataxin 表达的非病理水平,并使线粒体活动正常化。我们还优化了从健康和 FRDA 患者外周血中分离的 HSPC 中的基因编辑方法,并证明基因编辑细胞在体外和体内造血正常。该过程不会诱发细胞毒性作用或重大脱靶事件,但在基因编辑细胞中观察到 p53 介导的细胞增殖延迟。这项研究为将基因校正的 HSPC 自体移植用于 FRDA 的临床转化奠定了基础。