XiaoMi-AI文件搜索系统
World File Search SystemDNTPS

引文:Pietra D、Brisci A、Rumi E、Boggi S、Elena C、Pietrelli A、Bordoni R、Ferrari M、Passamonti F、De Bellis G、Cremonesi L 和 Cazzola M. Dee
对于 HRM 检测,采用补充表 S1 中报告的优化内含子引物。PCR 在 20 μ L 中进行,其中包含 100 ng DNA、0.5 单位 HotStart Taq 聚合酶以及 1x 缓冲液(Qiagen,德国希尔登)、1.5 mM MgCl 2、800 μ M dNTP、300 nM 每种引物和 1x EvaGreen(Idaho Technologies,犹他州盐湖城)作为插入染料。循环和 HRM 分析在 Rotor-Gene ™ 6000 实时分析仪上进行,采用以下热方案:95°C 持续 15 分钟(一个循环);95°C 持续 30 秒,55°C 持续 30 秒,72°C 持续 30 秒(50 个循环);72°C 持续 10 分钟(一个循环);熔化温度从 85°C 升至 95°C,每秒上升 0.1°C。使用相关的 Rotor-Gene ™ 6000 系列软件 (v1.7.87) 分析数据。标准化条在前导范围的 88°C 和 88.5°C 之间,在尾随范围的 92.5°C 和 93°C 之间,置信阈值为 90%:如果 HRM 图超出了指定参考基因型的置信范围,则软件会将样本识别为变异。图 S1A 显示了健康受试者和 3 名 MPN 患者的 DNA 样本的 HRM 图谱,这些样本先前已通过微电子微芯片分析进行了基因分型(未显示数据)。患者 PV02_113 为 MPL (W515K) 纯合子(TGG>AAG 转换),其 HRM 曲线相对于野生型序列向左移向较低温度,这与纯合变体导致熔解温度 (Tm) 降低的预期一致。患者 PV04_494 为 MPL (W515A) 纯合子(TGG>GCG 转换)等位基因

原始人的自然经济
焦磷酸测序:Roche模板由EMPCR 1制备,其中1-20万珠沉积在PTP井中。较小的珠,带有连接的硫酸酶和荧光素酶围绕模板珠。单个DNTP依次流过井,以预定的顺序分配。在掺入补体DNTP时,释放的PP I被转换为ATP,从荧光素蛋白到羟基二耐蛋白的氧化产生光。读取平均400个基础作为流程图。对于均聚物,重复多达六个核苷酸,添加的DNTP的数量与光信号成正比。插入是最常见的错误类型,其次是删除。通过连接测序:将约1亿个EMPCR的模板珠沉积在载玻片上。在退火时,添加了1,2个探针的库。适当的条件使选择性杂交和探针结扎到互补位置。1,2探针的第一个(y)和第二(z)位置被设计为审讯库,因此16个二核苷酸由四种染料编码。在四色成像之后,将带状的1,2探针化学裂解以产生5'-PO 4组(P)。杂交,连接,成像和裂解的循环又重复了六次。然后从模板中剥离扩展引物,并使用N – 1底漆进行第二个连接弹,该底漆将询问底座重置为左侧的一个位置。询问每个基础两倍,提高了颜色调用的准确性。随后发生了七个连接周期,然后再进行三个结扎弹。然后将35个数据位组成的字符串在色彩空间中编码,然后对准参考基因组以解码DNA序列。替换是最常见的错误类型。可逆终结器:DNA片段的Illumina Bridge放大是在载玻片的八个通道上随机分布的,高密度向前和反向引物共价附加到其上。固相扩增可从单个ssDNA模板产生约8000万个MC。将底漆退火到每个MC中模板的自由末端。聚合酶延伸,然后终止从四个RTs组中的DNA合成,每组用不同的染料标记。未合并的RT被洗净,通过四颜色成像进行基础识别,并通过化学裂解去除阻塞和染料组以允许下一个周期。给定MC的颜色图像提供了〜45个基础的读取。替换是最常见的错误类型。使用RTS进行单分子测序:Helicos数十亿个未夸大的ssDNA模板是用poly(da)尾巴制备的,这些尾巴与聚(DT)引物杂交,共同连接到载玻片上。对于一通测序,该引物 - 模板复合物就足够了。两通序测序涉及复制模板链,删除原始模板,并退火向表面(未显示)。与Illumina的RT不同,这四个Helicos RT用相同的染料标记,并以预定的顺序单独分配。融合事件导致荧光信号。使用单分子消除了Dephasing的问题,其中给定MC内的数千个复制模板不会有效地扩展其引物。删除是最常见的误差类型,可以通过提供约25个基本共识读取的两次测序可大大降低。的应用和挑战100篇论文描述了这些创新的成果。虽然改进继续,但读取长度限制,错误类型和频率显着影响组装策略。对于简短(<100个基本)读取平台,通过映射到参考基因组来指导组装。结合Sanger和Roche数据(100个基本读数)改善了从头组件2,并且随着焦磷酸测序读取长度的改进,使用混合Roche(250键读数)和Illumina数据进行了改善,已经描述了从头组装。最近使用Roche 4和Illumina平台报告了第一个个性化基因组测序项目。Roche,Illumina和AB平台在1,000个基因组项目中被用于生成人类遗传变异的详细图表以及人类微生物组项目,以将微生物组动态与人类健康相关联。应用不限于测序基因组。共识计数分析5最近出现了,从而实现了转录因子结合,mRNA剪接,DNA甲基化,小RNA,染色质结构和DNase超敏位点的全局分析。配对的测序方案。这些不仅对从头组件很重要,而且对于识别结构变化和映射mRNA剪接同工型。展望未来,太平洋生物科学,多佛系统(Polonator G.007),Visigen Biotechnologies,Lasergen,Inc。,Intelligent Bio-Symys,完整的基因组学和牛津Nanopore技术等公司的平台开发。
mytaqtm DNA聚合酶
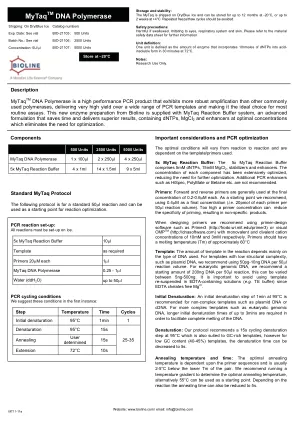
重要的考虑因素和PCR优化,最佳条件将从反应到反应,并取决于所使用的模板/引物。5x mytaq反应缓冲液:5x MyTAQ反应缓冲液包括5mm DNTP,15mm MGCL 2,稳定器和增强剂。每个组件的浓度已得到广泛优化,从而减少了进一步优化的需求。其他PCR增强剂,例如HISPEC,Polymate或Betaine等。。引物:正向和反向引物通常以0.2-0.6M的最终浓度使用。我们建议使用0.4M作为最终浓度(即每50 le反应体积的每个引物的下午20点)。 过高的底漆浓度可以降低启动的特异性,从而导致非特异性产物。 设计底漆时,我们建议使用Primer-Design软件,例如Primer3(http://frodo.wi.mit.edu/primer3)或可单位的10mm和3mm和3mm的阳离子阳离子浓度,或者单位阳离子浓度和分别为10mm和3mm。 引物应具有约60°C模板的熔化温度(TM):反应中的模板量主要取决于所使用的DNA类型。 对于低结构复杂性(例如质粒DNA)的模板,我们建议使用50pg-10ng DNA每50°L反应体积。 对于真核基因组DNA,我们建议每50 l反应的起始量为200ng DNA,这可以在5ng-5ng-500ng之间变化。 避免在含EDTA的解决方案中重新悬浮模板(例如)很重要 te buffer)由于EDTA螯合免费Mg 2+。每50 le反应体积的每个引物的下午20点)。过高的底漆浓度可以降低启动的特异性,从而导致非特异性产物。设计底漆时,我们建议使用Primer-Design软件,例如Primer3(http://frodo.wi.mit.edu/primer3)或可单位的10mm和3mm和3mm的阳离子阳离子浓度,或者单位阳离子浓度和分别为10mm和3mm。引物应具有约60°C模板的熔化温度(TM):反应中的模板量主要取决于所使用的DNA类型。对于低结构复杂性(例如质粒DNA)的模板,我们建议使用50pg-10ng DNA每50°L反应体积。对于真核基因组DNA,我们建议每50 l反应的起始量为200ng DNA,这可以在5ng-5ng-500ng之间变化。避免在含EDTA的解决方案中重新悬浮模板(例如te buffer)由于EDTA螯合免费Mg 2+。初始变性:对于非复合模板,例如质粒DNA或cDNA,建议在95°C下1分钟的初始变性步骤。对于更复杂的模板,例如真核基因组DNA,为了促进DNA的完全融化,需要更长的初始变性时间至3分钟。变性:我们的协议建议在95°C下进行15S循环变性步骤,这也适用于富含GC的模板,但是对于低GC含量(40-45%)模板,可以将变性时间降低到5s。退火温度和时间:最佳退火温度取决于底漆序列,通常比对下的TM低2-5°C。我们建议运行温度梯度以确定最佳退火温度,另外55°C可以用作起点。取决于反应,退火时间也可以减少到5s。

全基因组测序和分析 - Illumina
全基因组测序和分析 - 基于 Illumina Rhabdoid (RT) Illumina 基因组板的文库构建(350-450bp 插入大小):将 96 孔格式的 2ug 基因组 DNA 通过 Covaris E210 超声处理 30 秒进行碎裂,使用 20% 的“占空比”和 5 的“强度”。双端测序文库是按照 BC 癌症机构基因组科学中心 96 孔基因组 ~350bp-450bp 插入 Illumina 文库构建协议在 Biomek FX 机器人(Beckman-Coulter,美国)上准备的。简单来说,DNA在96孔微量滴定板中用Ampure XP SPRI 珠子纯化(每60uL DNA 40-45uL 珠子),在单一反应中分别用T4 DNA聚合酶、Klenow DNA聚合酶和T4多核苷酸激酶进行末端修复和磷酸化,然后用Ampure XP SPRI 珠子进行清理,并用Klenow片段(3'到5'外显子减去)进行3' A加尾。用Ampure XP SPRI 珠子清理后,进行picogreen定量以确定下一步接头连接反应中使用的Illumina PE接头的数量。使用 Ampure XP SPRI 珠子纯化接头连接产物,然后使用 Illumina 的 PE 索引引物组,用 Phusion DNA 聚合酶(美国赛默飞世尔科技公司)进行 PCR 扩增,循环条件为:98˚C 30 秒,然后 6 个循环,98˚C 15 秒,62˚C 30 秒,72˚C 30 秒,最后在 72˚C 延伸 5 分钟。使用 Ampure XP SPRI 珠子纯化 PCR 产物,并使用高灵敏度分析(美国珀金埃尔默公司)用 Caliper LabChip GX 检查 DNA 样本。所需大小范围的 PCR 产物经过凝胶纯化(在内部定制机器人中使用 8% PAGE 或 1.5% Metaphor 琼脂糖),并使用 Agilent DNA 1000 系列 II 检测和 Quant-iT dsDNA HS 检测试剂盒使用 Qubit 荧光计(Invitrogen)评估和量化 DNA 质量,然后稀释至 8nM。在使用 v3 化学法在 Illumina HiSeq 2000/2500 平台上生成 100bp 配对末端读数之前,通过 Quant-iT dsDNA HS 检测确认最终浓度。全基因组亚硫酸盐-Seq 文库构建和测序:使用 1-5 mg Qubit(Life Technologies,加利福尼亚州卡尔斯巴德)定量基因组 DNA 进行文库构建,如所述(Gascard 等人,2015 年)。为了追踪亚硫酸盐转化的效率,将 1 ng 未甲基化的 lambda DNA (Promega) 掺入使用 Qubit 荧光法定量的 1 µg 基因组 DNA 中,并排列在 96 孔微量滴定板中。使用 Covaris 超声处理将 DNA 剪切至 300 bp 的目标大小,并使用 DNA 连接酶和 dNTP 在 30o C 下对片段进行末端修复 30 分钟。使用 2:1 AMPure XP 珠子与样品比例纯化修复后的 DNA,并在 40 µL 洗脱缓冲液中洗脱以准备 A 尾;这涉及使用 Klenow 片段和 dATP 将腺苷添加到 DNA 片段的 3' 端,然后在 37o C 下孵育 30 分钟。用磁珠清理反应后,将胞嘧啶甲基化双端接头(5'-AmCAmCTmCTTTmCmCmCTAmCAmCGAmCGmCTmCTTmCmCGATmCT-3' 和 3'-GAGmCmCGTAAGGAmCGAmCTTGGmCGAGAAGGmCTAG-5')在 30oC 下连接到 DNA 20 分钟,并纯化接头两侧的 DNA 片段珠。在亚硫酸盐转化之前,用 10 个 PCR 循环扩增一份文库片段,并在 Agilent Bioanalyzer 高灵敏度 DNA 芯片上进行大小测定。扩增子的长度在 200-700 bp 之间。使用 EZ Methylation-Gold 试剂盒(Zymo Research)按照制造商的方案实现甲基化接头连接的 DNA 片段的亚硫酸盐转化。五次循环