XiaoMi-AI文件搜索系统
World File Search SystemDSBS

抗血管病原体的水力发电
。cc-by-nc 4.0国际许可证(未经同行评审证明)获得的是作者/资助者,他授予Biorxiv授予Biorxiv的许可,以永久显示预印本。这是该版本的版权所有,该版本发布于2024年7月5日。 https://doi.org/10.1101/2021.10.21.465387 doi:biorxiv Preprint


(缺少)DNA双链断休息途径在V(d)J重组期间选择
DNA双链断裂(DSB)是可以通过多种DNA修复途径修复的剧毒病变。多个因素可能会影响修复对给定途径的选择和限制,以保证维持基因组完整性。在V(D)J重组期间,RAG诱导的DSB(几乎)是通过非同理端连接(NHEJ)途径仅修复的,以实现抗原受体基因多样性的益处。在这里,我们回顾了将RAG生成的DSB修复到NHEJ的各种参数,包括RAG核酸酶产生的DNA DSB末端的特殊性,裂解后突触复合物的建立和维护,以及DNA末端的DNA末端的末端抗切除和(Microtro)的人体学修复。在这种生理背景下,我们强调某些DSB的DNA修复途径选择有限。

1介导减数分裂双链破裂活动
在第一个减数分裂细胞分裂中摘要,大多数生物体的染色体的适当分离取决于chiasmata,这是源自spo11核酸酶催化的编程双链断裂(DSB)的同源染色体之间的连续性交换。由于DSB会导致生殖细胞无法弥补的损害,而缺乏DSB的染色体也缺乏Chiasmata,因此必须仔细调节DSB的数量既不会太高也不太低。在这里,我们表明,在秀丽隐杆线虫中,减数分裂DSB水平受DSB-1的磷酸调节控制,DSB-1是PP4 PPH-4.1磷酸酶和ATR ATL-1 Kinase的相对活性,DSB-1(酵母SPO11辅助剂REC114)的同源物。PPH-4.1突变体中DSB-1磷酸化的增加与DSB形成的减少相关,而DSB-1磷酸化的预防大大增加了PPH-4.1突变体和野生型背景中的减数分裂DSB的数量。秀丽隐杆线虫及其近亲还具有DSB-1的差异旁系同源物,称为DSB-2,而DSB-2的丢失却可以减少年龄增加的卵母细胞中的DSB形成。我们表明,DSB-1的哲学和灭活形式的比例随着年龄的增长和DSB-2的流失而增加,而不可磷酸化的DSB-1则挽救了DSB-2突变体中DSB的年龄依赖性降低。这些结果表明,DSB-2部分进化以补偿DSB-1通过磷酸化的失活,以维持老年动物的DSB水平。我们的工作表明,PP4 PPH-4.1,ATR ATL-1和DSB-2与DSB-1协同作用,以在整个生殖寿命中促进最佳DSB水平。

科学期刊
泛素化是通过电离辐射(IR)诱导的DNA双链断裂(DSB)的正确修复所需的至关重要的翻译后修饰。dsbs主要通过同源重组(HR)修复,并且在不存在的情况下非同源末端连接(NHEJ)。此外,微型学介导的终端连接(MMEJ)和单链退火(SSA)提供了备份DSBS修复途径。然而,控制其使用的机制仍然知之甚少。通过在IR之后使用泛素系统的高分辨率CRISPR筛选,我们会系统地揭示细胞存活所需的基因,并阐明E3泛素连接酶SCF Cyclin F在依赖细胞周期依赖性DSB修复中的关键作用。我们表明,SCF细胞周期蛋白介导的EXO1降解可防止有丝分裂中的DNA末端切除,从而允许MMEJ发生。此外,我们确定了一个保守的细胞周期蛋白识别基序,与其他细胞周期蛋白所使用的基序不同,对细胞周期蛋白的特异性具有广泛的影响。

通过DNA聚合酶辅助的终端标记
在彗星测定中的摘要中,如果细胞被X X倍化为Genoto XIC剂,则在单细胞凝胶电泳后形成尾巴。these尾巴包括DNA单链断裂(SSB)和双链断裂(DSB)的混合物。ho w e v er,这些两种类型的链断裂无法使用具有Con V en ventionDNA染色的彗星测定方案来区分。由于DSB对单元格是有问题的,因此如果可以在同一彗星中差异化SSB和DSB,则将很有用。为了能够区分SSB和DSB,我们为聚合酶辅助的DNA损伤分析(PADDA)设计了一种协议,可与Flash Comet协议或固定单元格结合使用。通过使用DNA聚合酶I将SSB和末端脱氧核苷酸转移酶标记为具有荧光团标记的核苷酸的DSB。在此,TK6细胞或HACAT细胞暴露于过氧化氢(H 2 O 2),电离辐射(X射线)或DNA切割酶,然后遵循彗星方案,以实施彗星方案。p adda提供了更广泛的检测范围,未发现的DNA链断裂的未发现的未发现。
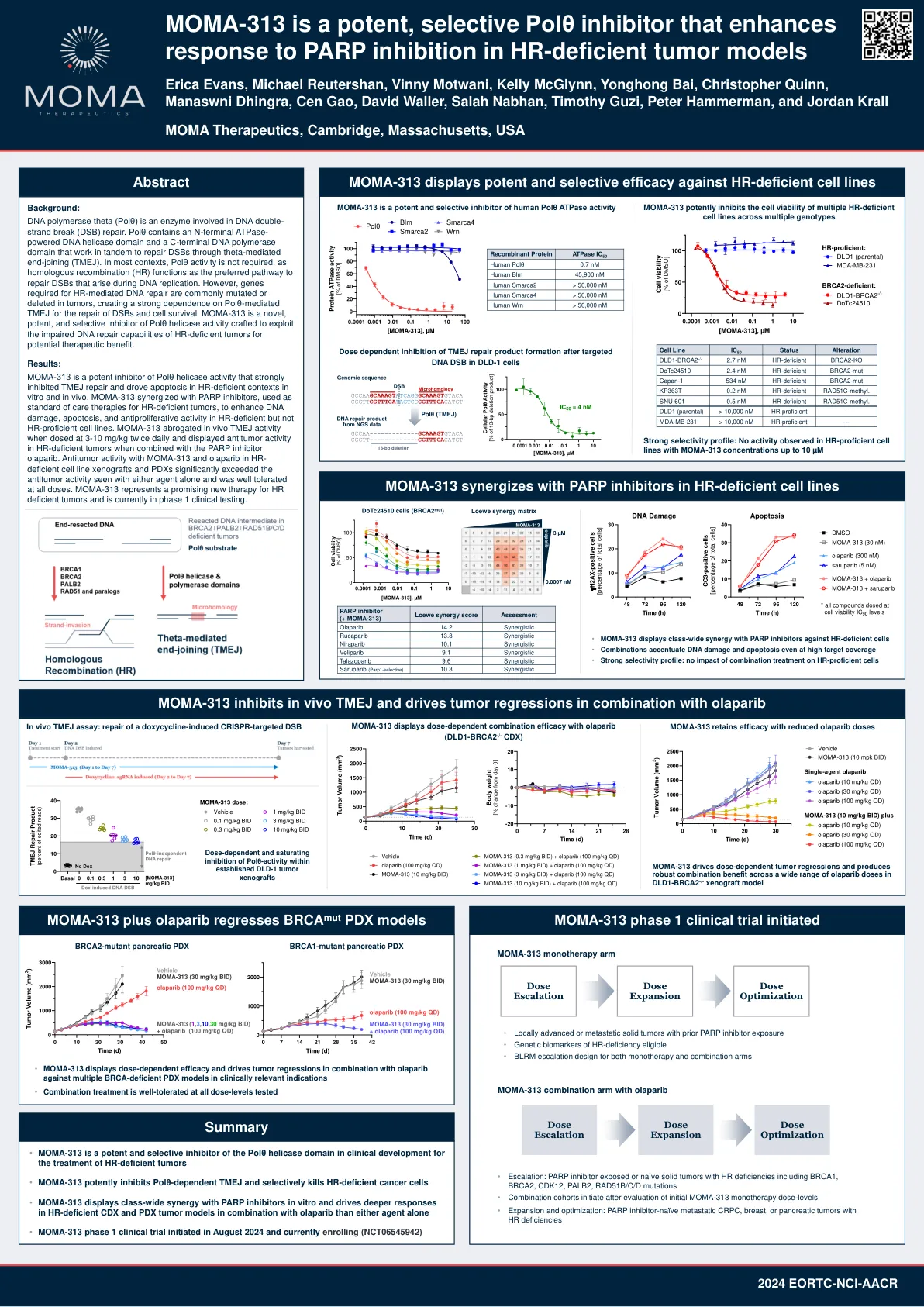
MOMA-313 是一种强效、选择性 Polθ 抑制剂,可增强 HR 缺陷肿瘤模型对 PARP 抑制的反应
DNA 聚合酶 theta (Polθ) 是一种参与 DNA 双链断裂 (DSB) 修复的酶。Polθ 包含一个 N 端 ATPase 驱动的 DNA 解旋酶结构域和一个 C 端 DNA 聚合酶结构域,它们协同作用,通过 theta 介导的末端连接 (TMEJ) 修复 DSB。在大多数情况下,Polθ 活性不是必需的,因为同源重组 (HR) 是修复 DNA 复制过程中出现的 DSB 的首选途径。然而,HR 介导的 DNA 修复所需的基因通常在肿瘤中发生突变或缺失,导致 DSB 修复和细胞存活严重依赖 Polθ 介导的 TMEJ。MOMA-313 是一种新型、有效且选择性的 Polθ 解旋酶活性抑制剂,旨在利用 HR 缺陷型肿瘤受损的 DNA 修复能力来获得潜在的治疗益处。

丁酰乳糖A是一种磷脂氟脂肪抑制剂,可增强
修复DNA损伤对于所有生物体来说都是至关重要的。DNA双链断裂(DSB)是最严重的DNA损伤类型之一,因为它们导致丧失了网络信息和未修复时死亡。在大肠杆菌中,它们被RECBCD复合物认识和处理,该复合物通过同源重组启动修复。尽管RECBCD下游的重复动力学已得到很好的特征,但尚不清楚该复合物与DNA保持附着多长时间,以及什么触发了其在体内的分离。要回答这些问题,我们在单分子水平上成像了RECB,并量化了其在暴露于环丙沙星的细菌细胞中的动态行为,这是一种诱导DSB的抗生素。我们的结果表明,RECB与DSB(10秒)形成长寿命的复合物,并且其与DNA的解离是复合物的固有证券,不取决于DNA损伤的量,也不取决于修复途径中的以下步骤。更重要的是,我们表明我们可以使用与DSB的RECB结合作为估计损害形成速率的标记。这项研究对RECBCD与DNA双链在体内的大肠杆菌的相互作用以及对环丙沙星诱导的DSB的细菌反应提供了详细的定量见解。

科学期刊
体细胞中双链断裂 (DSB) 的修复主要通过易出错的非同源末端连接完成,较少通过精确的同源定向修复完成,优先使用姐妹染色单体作为模板。在这里,果蝇系统使用同源染色体的完整序列对 DSB 和单链断裂 (SSB) 进行有效的体细胞修复,我们称这一过程为同源染色体模板修复 (HTR)。出乎意料的是,白色位点的 HTR 介导的等位基因转换对 Cas9 衍生的切口酶 D10A 或 H840A 诱导的 SSB 的响应比对完全活性 Cas9 诱导的 DSB 的响应 (20% 到 30%) 更有效 (40% 到 65%)。 Nickase 和 Cas9 引起的修复表型在发展时间(分别为晚期和早期)和不良诱变事件的产生(罕见和频繁)方面均有所不同。Nickase 介导的 HTR 代表了一种高效且出乎意料的等位基因校正机制,在基因编辑领域具有深远的潜在应用。
将核酸外切酶与 Cas9 融合可增强毕赤酵母中的同源重组
HR 比 NHEJ 慢得多,NHEJ 可以从 DSB 事件中拯救更多细胞。NHEJ 几乎不需要或根本不需要末端切除来直接重新连接 DSB 末端。相比之下,HR 需要短距离切除和长距离切除 DSB 以及供体来实施修复过程。此外,其他蛋白质也可能是 HR 修复途径的限制因素 [18, 19]。我们在此发现,在同时删除两个基因和整合多个片段期间,将 MRE11 与 CAS9 融合可提高 CFU 数量