XiaoMi-AI文件搜索系统
World File Search SystemEvolutionary


进化计算
SCS5107 计算智能 计算智能:计算智能 (CI) 是一套受自然启发的计算方法和方法,用于解决现实世界应用中的复杂问题,而传统方法和方法对此无效或不可行。它主要包括模糊逻辑系统、神经网络和进化计算。此外,CI 还包含源自上述三种技术或围绕其中一种或多种技术的技术,例如群体智能和人工免疫系统,它们可以看作是进化计算的一部分。进化计算在计算机科学中,进化计算是人工智能(更具体地说是计算智能)的一个子领域,涉及组合优化问题。进化技术主要涉及元启发式优化算法,例如:进化算法(包括遗传算法、进化规划、进化策略和遗传规划)群体智能(包括蚁群优化和粒子群优化,以及较小程度的人工免疫系统、文化算法、差异进化、和谐搜索算法等。在人工智能中,进化算法 (EA) 是进化计算的一个子集,是一种基于种群的通用元启发式优化算法。EA 使用一些受生物进化启发的机制:繁殖、突变、重组和选择。优化问题的候选解决方案扮演着种群中个体的角色,适应度函数决定了解决方案“生存”的环境(另见成本函数)。在重复应用上述运算符后,种群就会进化。群体智能在计算智能领域有两种流行的群体启发方法:- 蚁群优化 (ACO)

动态进化方法...
多任务学习假设能够从多个任务中学习的模型可以通过知识迁移实现更好的质量和效率,这是人类学习的一个关键特征。然而,最先进的 ML 模型依赖于每个任务的高度定制,并利用大小和数据规模而不是扩展任务数量。此外,持续学习将时间方面添加到多任务中,通常专注于研究常见的陷阱,例如灾难性遗忘,而不是将其作为构建下一代人工智能的关键组成部分进行大规模研究。我们提出了一种能够生成支持动态添加新任务的大规模多任务模型的进化方法。生成的多任务模型是稀疏激活的,并集成了基于任务的路由,可保证在模型扩展时计算成本有限并且每个任务添加的参数更少。所提出的方法依赖于知识分区技术来实现对灾难性遗忘和其他常见陷阱(如梯度干扰和负迁移)的免疫。我们通过实验证明,所提出的方法可以联合解决 69 个公共图像分类任务并取得有竞争力的结果,例如,与在公共数据上训练的最佳模型相比,通过实现 15% 的相对误差减少,提高了 cifar10 等竞争基准的最新水平。


螯肢动物的进化生物学
螯肢动物门是一类古老、生物多样性丰富且生态意义重大的节肢动物。过去十年,螯肢动物进化研究经历了一次复兴,使我们对高级系统发育和生物目内部关系的理解发生了重大变化。这些概念上的进步包括在螯肢动物目子集中发现多个全基因组复制事件,例如马蹄蟹、蜘蛛和蝎子。因此,螯肢动物进化的长期假设和教科书场景,例如蛛形纲的单系性和蛛形纲共同祖先的一次陆地殖民,引起了争议。该谱系中古老的重复基因的保留也为研究基因复制在螯肢动物宏观进化中的作用提供了沃土。这一新的研究前沿与第一种针对蛛形纲动物模型的基因编辑协议的及时建立相同步,促进了新一代实验方法的出现。
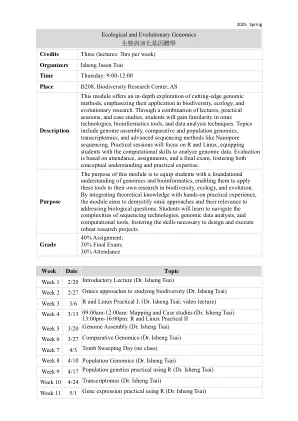
进化生物学家的生物信息学
本独立的教科书涵盖了进化生物学中序列分析的基本方面,包括序列比对,系统发育重建和融合模拟。它通过一系列超过400个计算机问题来解决这些方面,从基础到研究级别,再到完成学习。学生在科学数十年的相同计算环境中解决了问题 - UNIX命令行。这在PC的所有三个主要操作系统上都可用:Microsoft Windows,Mac-OSX和Linux。要使用此功能强大的系统学习,学生通过应用通用工具,生物信息学软件以及专门为本课程编写的40多个程序来分析样本序列数据。包括所有问题的解决方案,这本书是自学的理想之选。问题分为以引言和新概念和程序列表为首的部分。通过使用实用计算来探索进化概念和序列数据,该书使读者能够解决自己的计算问题。

(进化)变革之风:
这种二分法的问题和有害性在于,原核生物最初在细胞学上被定义为负面的。换句话说,原核生物缺乏真核细胞的这种或那种特征:甚至油滴或凝聚层都符合这种负面定义。原核生物-真核生物二分法的任何优点在于它有助于理解真核生物,而真核生物可能是通过“原核”阶段进化而来的。随着重复(作为教义问答),原核生物-真核生物二分法只会让微生物学家轻易接受他们对原核生物之间关系几乎一无所知的事实;他们甚至对这一事实——当今最大的挑战之一——感到迟钝,即他们丝毫不了解原核生物和真核生物之间的关系。细菌之间的关系问题归结为“如果它不是真核生物,而是原核生物”,而要了解原核生物,我们只需确定大肠杆菌与真核生物有何不同。这并不是对创造性思维的邀请,也不是统一的生物学原理。这种真核生物-原核生物二分法是原核微生物学与真核微生物学之间的一道障碍。这种对微生物学的短视观点不仅未能认识到微生物关系问题的重要性,而且未能认识到今天难以解决的问题明天可能并非如此。自 20 世纪 50 年代以来,分子序列就被用于确定进化关系,而 Zuckerkandl 和 Pauling 的开创性文章“分子作为进化历史的记录”在 1965 年最令人信服地阐述了这一观点(36)。然而,记录表明,微生物学——最需要的生物科学——实际上对这些方法的意义和潜力视而不见。然而,在 20 世纪 70 年代末,情况发生了巨大变化。rRNA 序列已被证明是原核生物系统发育的关键(例如 8)。尽管原核生物在细胞和生理水平上没有提供可靠的系统发育排序特征,但它们的 rRNA 足以做到这一点。到 20 世纪 80 年代初,随着基于 rRNA 的原核生物系统发育开始出现,微生物学家开始(尽管非常缓慢地)重新意识到了解微生物系统发育的重要性。将所有原核生物视为同一种类的愚蠢做法,在古细菌(最初称为古细菌)的发现中得到了戏剧性的揭示。古细菌是一类完全出乎意料的原核生物,如果真要说有什么不同的话,那就是它与真核生物(真核生物)的关系比与其他原核生物(真正的)细菌(11、13、32、34)的关系更密切。即便如此,真核生物的力量——

进化计算手册
《进化计算手册》是进化计算 (EC) 领域的一个重要里程碑。与任何新领域一样,进化计算也经历了多个不同的发展和成熟阶段。该领域始于 20 世纪 50 年代末和 60 年代初,当时数字计算的出现使得科学家和工程师能够构建和试验各种进化过程模型。这项早期工作产生了许多重要的 EC 范式,包括进化规划 (EP)、进化策略 (ES) 和遗传算法 (GA),这些范式成为 20 世纪 70 年代大部分工作的基础,这一时期人们对这些思想进行了深入的探索和改进。其结果是产生了各种强大的算法,它们具有解决困难的科学和工程问题的巨大潜力。到 20 世纪 80 年代末和 90 年代初,活动水平已经发展到与主要 EC 范式(GA、ES 和 EP)相关的每个子组都参与规划和举办自己的定期会议的程度。然而,在该领域内,人们越来越意识到需要加强各个子组之间的互动和凝聚力。如果整个领域要成熟,它需要一个名称,它需要有一个清晰的凝聚力结构,它需要一个档案文献库。20 世纪 90 年代反映了这种成熟,选择进化计算作为该领域的名称,建立了两个期刊

进化增强学习
摘要 - 在复杂解决问题中的增强学习和进化算法(EAS)的局限性时,进化增强学习(EVORL)已成为协同解决方案。Evorl整合了EAS和增强学习,为培训智能代理提供了有希望的途径。该系统评价首先通过Evorl的技术背景导航,研究了EAS和强化学习算法之间的共生关系。然后,我们深入研究了EAS和强化学习所面临的挑战,探索了它们的相互作用和对Evorl功效的影响。此外,审查强调了解决与Evorl当前景观内有关可伸缩性,适应性,样本效率,对抗性鲁棒性,道德和公平性有关的开放性问题的必要性。最后,我们提出了Evorl的未来方向,强调了努力增强自我适应和自我改善,概括,可解释性,可解释性等的研究途径。作为研究人员和从业人员的综合资源,这项系统的审查提供了有关Evorl现状的见解,并提供了指南,以推动其在不断发展的人工智能景观中的能力。