XiaoMi-AI文件搜索系统
World File Search SystemSH2

可离子网络介导SH2信号蛋白中的pH依赖性变构
对于具有生理相关的预测PK A值的可离子残基,并且数据在3D结构或2D残基相互作用网络中可视化。(b)以卡通和表面格式显示的SHP2的晶体结构(PDB ID:2SHP)。蛋白质酪氨酸磷酸酶(PTP)结构域以灰色为灰色的SH2域颜色为黄色。(c)灰色和SH2结构域的SHP2(PDB ID:2SHP)的结构(PDB ID:2SHP)在黄色的灰色和SH2结构域中的结构。通过在球体中显示的可离子网络预测管道中通过的残基。带有预测PK A位移(青色)簇的残基,具有可离子相互作用的人(洋红色)跨磷酸酶-SH2域相互作用界面的残基。(d)在47 SHP2结构(平均值±SD)上使用硅离子化网络预测管道鉴定出的青色残基的预测PK A S的表。(e)残基的残基相互作用网络具有预测的PK A Shifts(Cyan)及其可电离相互作用器(Magenta)。边缘的长度反映了库仑相互作用的强度,在PTP-SH2相互作用界面处,较强的库仑相互作用具有更短的边缘长度(F)SHP2结构的变焦。来自A和B的网络残基显示在棒子中。残基有预测的PK a在青色和洋红色中的电离相互作用者的变化。

一些化合物的合成、表征及抗真菌活性
近二十年来,过渡金属配位化合物由于其独特性质(如催化、离子交换、微电子、非线性光学、多孔材料等)的合成及应用已成为一个极具吸引力的领域。[1-7] 过渡金属混合配体配合物在光化学、分析化学和磁化学等不同领域发挥着重要作用。[8] 锰的配位化学已成为生物无机化学中一个令人感兴趣的研究领域。[9] 目前,人们正在探索此类化合物的磁性和多种催化活性,以了解其生物学重要性。[10-12] +3 氧化态的锰 (Mn) 与带电和中性配体形成复合物。[13] 我们给出了实验室合成的三(乙酰丙酮)-锰 (III) 单晶的 X 射线晶体学数据。[14] 已发现锰 (III) 八面体配合物易受 Jahn-Teller 畸变的影响。我们进一步合成了四种新型混合配体 Mn(III) 配合物,即 [Mn(acac)2(NCS)SH2]、[Mn(acac)2(N3)SH2]、[Mn(acac)2(Cl)SH2] 和 [Mn(acac)2(Br)SH2],并研究了它们的磁化率、紫外线和抗真菌性能。



对“无药可用”的致病蛋白质进行药物治疗:重点...
信号转导和转录激活因子 (STAT) 3 被归类为“不可用药”的致病蛋白,尽管它含有 Src 同源性 (SH) 2 结构域,这是一个潜在的致命弱点,过去 30 年来一直未能被学术界和制药界成功针对。基于突变和建模研究,我们的小组开发了一种独特的虚拟配体筛选策略,针对 STAT3 SH2 结构域,结合强大的生化和细胞分析以及基于结构的药物化学,并鉴定出 TTI-101。TTI-101 是临床开发中最先进的直接小分子含 SH2 结构域致病蛋白抑制剂之一。TTI-101 目前正在进行 1 期研究,以确定其在晚期实体瘤患者中的安全性和耐受性以及药效学作用和疗效。
mRNA的表观遗传调节介导表型...
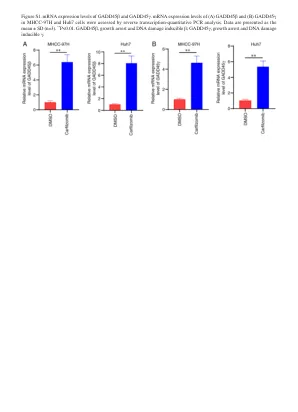
图S2。 通过蛋白质印迹评估的GADD45αshRNA的沉默效率。 gADD45α蛋白表达水平在(a)MHCC -97H和(b)用NC和三个靶向GADD45α的SHRNA后的HUH7细胞中。 基于GADD45α蛋白的表达,SH2的沉默是最重要的,用于随后的实验。 数据表示为平均值±SD(n = 3)。 ** p <0.01。 GADD45α,生长停滞和DNA损伤诱导α; NC,阴性对照; SH,短发夹。图S2。通过蛋白质印迹评估的GADD45αshRNA的沉默效率。gADD45α蛋白表达水平在(a)MHCC -97H和(b)用NC和三个靶向GADD45α的SHRNA后的HUH7细胞中。基于GADD45α蛋白的表达,SH2的沉默是最重要的,用于随后的实验。数据表示为平均值±SD(n = 3)。** p <0.01。GADD45α,生长停滞和DNA损伤诱导α; NC,阴性对照; SH,短发夹。

RASA1在癌症中的作用 EV20 −SSS − VC/MMAF,一种靶向抗体&drug ... 与顺铂,紫杉醇和... 的组合处理 潜在威胁生命的严重皮肤不良... 载脂蛋白负载的纳米颗粒抑制了生长,... IV期EGFR突变阳性NSCLC 的总体生存率 NLRP3炎性途径的抑制剂AS ... Rho相关激酶在肺癌中的作用(综述) 在... 中长期非编码RNA的机制和功能 低氧诱导因子2α在肺癌中的作用(综述) 当前的患者衍生肿瘤的研究发展... 内质网应激诱导的细胞死亡作为... TERT及其潜在治疗的肿瘤效应... 依维莫司治疗和副作用:系统评价... 肺癌患者的皮肤不良反应... 新辅助治疗的治疗模式和进度... 乳腺癌中的酪氨酸激酶抑制剂(审查) ... 的细胞穿透肽指导的最新应用 通过... 评估心脏剂量预测的准确性
RAS P21蛋白激活剂1(RASA1)位于铬-5q14.3上,是Rasgap家族的成员,其中包括NF1,DAB2IP和Rasal2(1)。RASA1包含以下域:SRC同源性2和3(SH2和SH3),N末端C2A和C2B,GTPase激活蛋白(GAP)和Pleckstrin同源(pH),它们附着在Bruton的酪氨酸酶(BTK)基础上。rasa1是具有双重指定性的差距,可增强和加速RAS和RAP的GTPase活性。值得注意的是,细胞内Ca 2+水平调节RASA1的间隙活性。当Ca 2+浓度较高时,RAS的C2结构域和RAP允许磷酸脂质的结合,而pH结构域则保持不活跃并防止脂质结合。rasa1通常位于细胞质中,作为可溶性蛋白质,并在细胞内Ca 2+浓度的受体介导的增加后募集到质膜上(2)。当RASA1与膜相关时,RASA1的RasGAP活性增加了,因为RasGap活性以RASA1的可溶形式有限,尽管未知的机制尚不清楚(3)。sh2 -ptyr相互作用允许RASA1与P190RHOGAP(P190RHOGAP -A,ARHGAP35)相互作用,这是Rho的差距(4)。由于其特殊
2024 年季风课程(UG1 除外)的讲座时间表
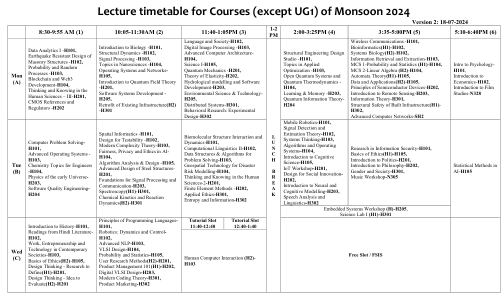
无线通信 - H101、生物信息学 (H1) - H102、系统生物学 (H2) - H102、信息检索与提取 - H103、MCS 1-概率与统计 (H1) - H104、MCS 2-线性代数 (H2) - H104、自动机理论 (H1) - H105、数据与应用 (H2) - H105、半导体器件原理 - H202、遥感简介 - H203、信息理论 - H301、建筑基础设施结构安全 (H1) - H302、高级计算机网络 - SH2

SOCS1与多种信号蛋白结合并抑制钢因子依赖性增殖
我们将SOCS1识别为试剂盒受体酪氨酸激酶信号通路的下游成分。我们表明,暴露于钢因子后,SOCS1 mRNA的表达迅速增加,而SOCS1通过其SRC同源性2(SH2)结构域与Kit受体酪氨酸激酶结合。先前的研究表明,SOCS1抑制了抑制Janus家族激酶的M1细胞的细胞因子介导的分化。相比之下,SOCS1的本构表达抑制了试剂盒的有线剂潜力,同时保持依赖钢因子的细胞存活信号。与Janus激酶不同,SOCS1不抑制KIT酪氨酸激酶的催化活性。为了定义SOCS1介导的抑制KIT依赖性有丝分裂发生的机制,我们证明SOCS1与信号蛋白GRB-2和Rho-family鸟嘌呤核苷酸交换因子VAV VAV结合。我们表明,GRB2通过其SH3结构域结合SOCS1与位于SOCS1 N末端的假定二苯胺决定因素,SOCS1与VAV的N末端调节区域结合。这些数据表明SOCS1是一个可诱导的开关,它调节增生信号,有利于细胞存活信号,并用作受体酪氨酸激酶信号途径中的衔接蛋白。关键字:grb2/sh2/sh3/信号转导/vav
多路复用 CRISPR-Cas12a 基因编辑的健康供体来源的 NK 细胞疗法,增加颗粒酶 B 和脱颗粒,支持改进的连续杀伤
SK-OV-3 肿瘤细胞 +10 ng/mL TGF-β 随时间变化,E:T 比例为 5:1;100 小时后测量肿瘤细胞。代表来自 n=2 名捐赠者的 3 次独立实验。误差线为标准偏差。CISH,细胞因子诱导的含 SH2 蛋白;E:T,效应物:靶标;GzmB,颗粒酶 B;NK,自然杀伤细胞;TGFBR2,转化生长因子 β 受体 II。