XiaoMi-AI文件搜索系统
World File Search SystemTCR
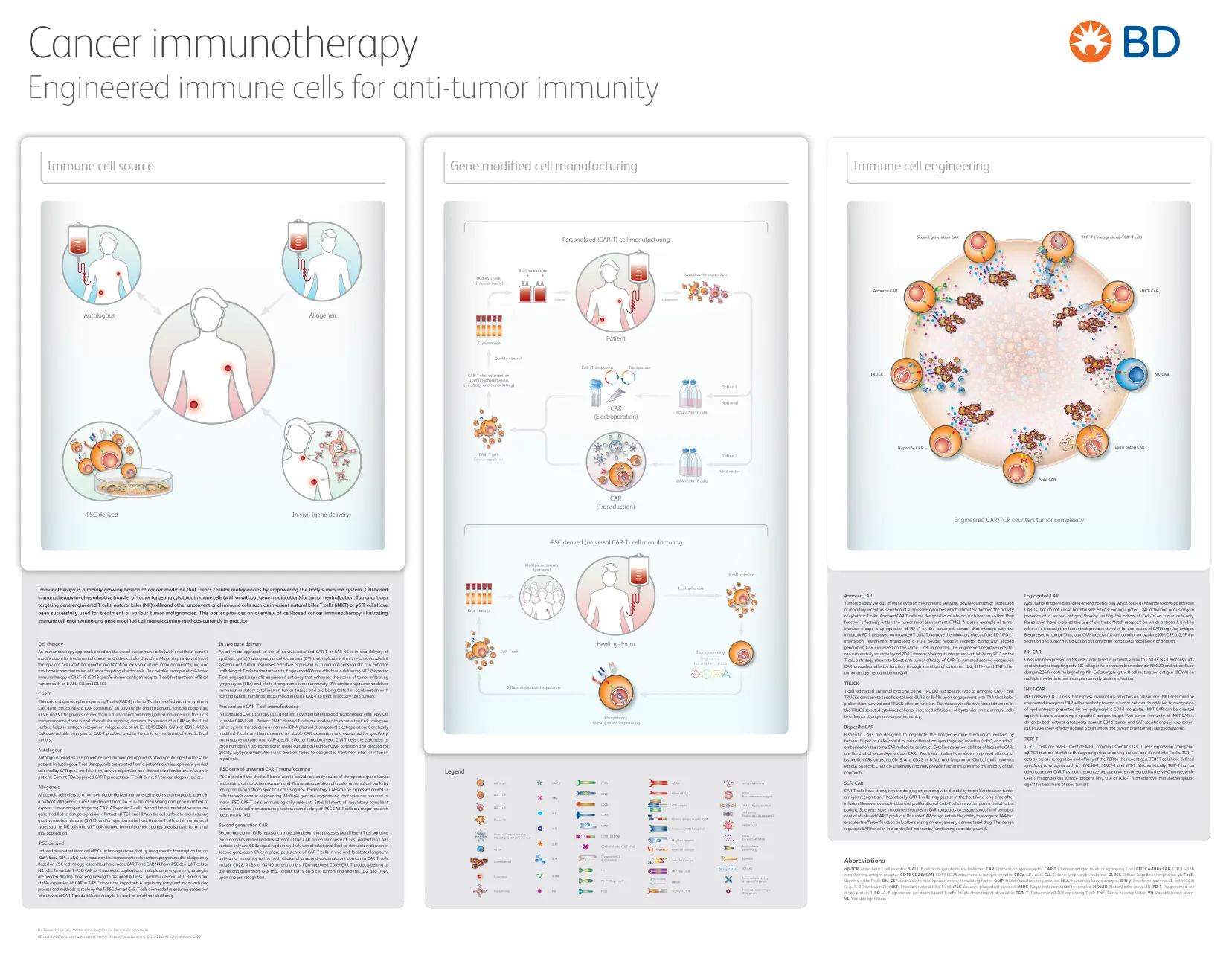
癌症免疫疗法海报-BD Biosciences
αβ-TCR,αβT细胞受体; B-所有,B细胞急性淋巴细胞白血病;汽车,嵌合抗原受体; CAR-T,表达T细胞的嵌合抗原受体; CD19 4-1BBZ汽车,CD19 4-1BB Zeta嵌合抗原受体; CD19 CD28Z汽车,CD19 CD28 Zeta嵌合抗原受体; CD3Z,CD3 Zeta; CLL,慢性淋巴细胞性白血病; DLBCL,使用大B细胞淋巴瘤; γδT细胞,伽马三角细胞; GM-CSF,粒细胞 - 巨噬细胞群刺激因子; GMP,良好的制造实践; HLA,人白细胞抗原; IFN-γ,干扰素伽玛; IL,白介素(例如IL-2白介素2); inkt,不变的天然杀手T细胞; IPSC,诱导多能干细胞; MHC,主要的组织相容性复合物; NKG2D,天然杀手级2d; PD-1,程序性细胞死亡蛋白1; PD-L1,程序性细胞死亡配体1; SCFV,单链片段变量; TCR + T,转基因αβ-TCR表达T细胞; TNF,肿瘤坏死因子; VH,可变重链; VL,可变轻链

T-DM1 耐药乳腺癌细胞的表征
图 1 MDA-MB-361 细胞系长期暴露于 T-DM1 会导致对 ADC 的敏感性降低。 (A) T-DM1 对 MDA-MB-361 S、TR 和 TCR 的 MTT 细胞毒性试验表明,与亲本相比,两种抗性细胞的 IC50 值均有所增加。 通过双向方差分析进行统计分析,然后进行 Bonferroni 后检验,并显示 TR (***:P < .001;**:P < .01;*:P < .05) 和 TCR (+) 与 S 相比的差异。 (B) 将亲本和抗性细胞暴露于浓度不断增加的 T-DM1 中,并使用 xCELLigence 跟踪细胞指数。 绘制了由 RTCA 软件确定的标准化细胞指数的斜率。统计分析采用双向方差分析,随后进行 Bonferroni 后检验,并显示每种细胞系在对照和暴露条件下的差异 (*: P < .05; ***: P < .001)。(C) 暴露于 T-DM1 6 天后,通过流式细胞术研究 Annexin 阳性细胞。与亲本细胞相比,TR 和 TCR 中的 Annexin 阳性细胞百分比有所下降。统计分析采用双向方差分析,随后进行 Bonferroni 后检验 (*: P < .05; ***: P < .001)

基于 RNP 的原代 CD4+T 细胞编辑...
电穿孔后 72 小时,可使用 BioLegend APC 抗人 TCR α / β 抗体通过流式细胞术评估用靶向 Edit-R sgRNA RNP 的 TRAC 或 TRBC 编辑的原代 CD4 + T 细胞的 TCR α / β 敲除情况。除了通过流式细胞术读取表型外,在基于 RNP 的编辑后 48-72 小时内,可以通过 T7EI/TIDE 测量插入/缺失形成。使用表 1 中列出的每个经过验证的 sgRNA 的引物,遵循 Dharmacon™ Edit-R™ 合成 gRNA 阳性对照试剂盒方案中的直接细胞裂解和 PCR 条件。要测量 T7EI 内切酶的插入/缺失形成,请完成上面列出的方案并使用分析软件。要通过分解 (TIDE) 分析跟踪插入/缺失来测量插入/缺失形成,请将得到的 PCR 扩增子发送至 Sanger 测序并使用网络工具,例如 http://shinyapps.datacurators.nl/tide/ 。以下方案描述了用于通过流式细胞术评估原代 CD4 + T 细胞中 TCR α / β 表型敲除的染色条件。1. 通过离心(300-5 分钟)沉淀用 PPIB、NTC2、TRAC 或 TRBC 靶向 RNP 电穿孔的 CD4 + T 细胞

CD3E(T细胞标记)抗体 簇蛋白 /载脂蛋白J(Apo-J)抗体< / div> alpha-1-抗胰蛋白酶(SERPINA3)(组织细胞瘤标记)抗体的产品图像 同源蛋白Nanog(癌症干细胞标记)抗体的产品图像 重组GFAP的产品图像(星形胶质细胞和神经干细胞标记)抗体 alpha-1-抗胰蛋白酶(SERPINA3)(组织细胞瘤标记)抗体的产品图像
特异性和评论识别CD3的Epsilon链,该链由MW的五个不同的多肽链(指定为Gamma,Delta,Epsilon,Zeta和Eta),MW为16-28KDA。CD3通常在高水平上表达在外周T细胞和大多数T细胞肿瘤上。胸腺细胞在分化过程中在细胞表面的不同水平上表达CD3,在皮质胸腺中,CD3主要是胞质内的。CD3复合物在淋巴细胞细胞表面与T细胞抗原受体(TCR)紧密相关,并参与将抗原识别信号转导到T细胞的细胞质中以及调节TCR复合物的细胞表面表达。

gmp 制造的 cas9 核酸酶细胞治疗 - ...
图 2. CTS Cas9 的 TCR 敲除效率高于供应商 A Cas9。将每个 Cas9 (7.5 pmol) 和靶向 alpha 和 beta T 细胞受体基因 (TRAC 和 TRBC) 区域的 Invitrogen ™ TrueGuide ™ 合成 sgRNA (7.5 pmol) 混合以创建 Cas9-RNP 复合物。每个 Cas9-RNP 复合物用于使用 Neon 转染系统 (货号 MPK5000) 转染 500,000 个 T 细胞。 72 小时后收获细胞,用 Invitrogen ™ eBioscience ™ 可固定活力染料 (FVD) 紫罗兰 (货号 65-0863-14) 和 eBioscience ™ 抗 TCR a/b 抗体 (货号 12- 9986-42) 染色,然后在 Invitrogen ™ Attune ™ NxT 流式细胞仪上进行分析,并使用基于 NGS 的 TAV 进行基因分型。 (A) TCR KO 效率的流式细胞术数据示例。与供应商 A Cas9 相比,CTS Cas9 的 KO 效率超过 88.7%,而供应商 A Cas9 的 KO 效率为 61.7%。 (B) 基于 NGS 的 TAV 的平均 KO 效率。与供应商 A Cas9 相比,CTS Cas9 在各种目标上实现了更高的平均 KO 效率。所有反应均重复进行三次 (** P < 0.01)。

HLA-A/B/C敲除电穿孔套件
说明HLA-A/B/C敲除电穿孔套件适用于通过电穿孔的细胞系和原代T细胞工程。该套件既包含Cas9酶(链球菌)和靶向HLA-A/B/C(人白细胞抗原)的GRNA。该套件足以设计高达500万个原代T细胞。背景HLA(人白细胞抗原)-a,b和c是MHC的三种主要类型(主要的组织相容性复合物)1类跨膜蛋白。它们与β2微球蛋白蛋白(由B2M基因编码)形成异二聚体。MHC 1类分子表现出短多肽,通常在长7-11个氨基酸之间,以识别为“自我”或“非自身”的免疫系统。HLA-C存在于所有细胞中,并且由于HLA-C基因的多样性而作为几种单倍型存在。c*08:02代表一种这样的单倍型。HLA I类将新抗原衍生的肽呈现到细胞表面,从而通过TCR(T细胞受体)识别出T细胞的识别。 癌症免疫疗法一直在使用该机制,方法是表达能够识别特定癌症免疫原子的TCR。 在2016年,HLA-C*08:02限制性TIL(肿瘤浸润淋巴细胞)在肺癌中靶向KRAS(Kirsten大鼠肉瘤病毒)G12D突变,导致阳性结果。 在转移性胰腺癌患者中采用了类似的方法,并导致该疾病的消退。 HLA-C*08:02限制性TIL对其他新抗原的TCR的研究可能对癌症治疗有益。 应用程序HLA I类将新抗原衍生的肽呈现到细胞表面,从而通过TCR(T细胞受体)识别出T细胞的识别。癌症免疫疗法一直在使用该机制,方法是表达能够识别特定癌症免疫原子的TCR。在2016年,HLA-C*08:02限制性TIL(肿瘤浸润淋巴细胞)在肺癌中靶向KRAS(Kirsten大鼠肉瘤病毒)G12D突变,导致阳性结果。在转移性胰腺癌患者中采用了类似的方法,并导致该疾病的消退。HLA-C*08:02限制性TIL对其他新抗原的TCR的研究可能对癌症治疗有益。应用程序K562细胞是HLA I和II类负的,使其成为引入和研究特定单倍型响应的理想细胞模型。hla在供体细胞和个体之间的不匹配可以导致免疫排斥反应,一种选择是敲除内源性HLA,从而使细胞被更广泛地普遍使用。

时间2 evolve:预测工程T细胞的功效
基因工程汽车和TCR转基因T细胞的抽象免疫疗法是癌症医学的一种变革性治疗方法。有一条具有靶抗原和先进技术的丰富管道,不仅可以在罕见的血液恶性肿瘤中,而且还可以在普通实体瘤中建立这种新颖的治疗方法。T2Evolve联盟是一种旨在加快癌症患者临床前开发和增加获得工程T细胞免疫疗法的临床前开发和访问权限的公共私人伙伴关系。T2Evolve中的一个关键野心是评估当前可用的临床前模型,以评估工程T细胞疗法的安全性和功效,并开发新模型和测试参数具有较高的临床安全性和功效预测价值,以改善和加速铅T-Cell产品的临床转换。在这里,我们回顾了允许评估汽车和TCR信号传导和抗原结合的现有和新兴临床前模型,工程T细胞对原发性和转移性肿瘤配体的访问和功能,以及内源性因素(例如宿主免疫系统和微生物组)的影响。总的来说,本评论文章介绍了基于创新的汽车和TCR转基因T细胞产品的创新标准化临床前测试系统的观点。

双特异性抗体重定向合成激动剂受体改性T细胞针对黑色素瘤
背景肉瘤是间充质细胞起源的一个异源性群体,通常很难以较差的预后治疗。大约30%的肉瘤的特征是表达充当致癌驱动因素的融合蛋白。脱落的小圆形细胞肿瘤(DSRCT)是由病原体EWSR1-WT1融合事件定义的典型融合驱动的肉瘤。所得的EWSR1-WT1致癌融合蛋白包含与正常自我蛋白不同的共有氨基酸序列。我们假设克隆保守的融合蛋白可能会产生共享或公共新抗原(NeoAgs)的免疫原性子集(NEOAGS),这可能是新型免疫治疗方法的靶标。使用HLA免疫沉淀/质谱法(HLA-IP/MS)屏幕的方法和结果,我们确定了从EWSR1- WT1融合蛋白的Junctim中得出的9-氨基酸肽序列(SsygQQS EK),均经常出现在HLA-WT1融合蛋白中,并在HLA-a*03中均出现。结合同源肽。我们证实了相同的肽序列在生理上由HLA-A*03 + DSRCT细胞呈现。使用带有MS识别的NEOAG的荧光团偶联的HLA-Multimers(Dextramers),我们检测到在HLA + DSRCT患者子集中结合融合NEOAG的循环T细胞,确认了免疫原性。随后,我们使用融合NEOAG特异性T细胞的体外抗原指导的克隆膨胀来隔离n = 3 HLA-A*03限制性,n = 1 HLA-A*11限制性融合NeoAg反应性克隆。使用单细胞测序,我们检索了由这些T细胞表达的T细胞受体(TCR)的TCRAB基因序列,并将其克隆到逆转录病毒表达载体中。多克隆CD8 + T细胞被检索到的TCR基因结合融合融合的葡萄晶剂,而不是载有病毒肽的控制葡萄糖剂。此外,表达候选TCR的CD8 + T细胞与表达必要的HLA等位基因和EWSR1-WT1融合的细胞稳健上调TNF A,并特别裂解HLA + DSRCT细胞。有趣的是,我们确定了一个独特的TCR,该TCR以肽范围的方式结合了融合NEOAG,该方式可以特异性地裂解HLA-A*03 +和-a*11 + dsRCT细胞,但没有在HLA-A*02的上下文中使用融合NeoAg。这意味着单一的TCR治疗可能覆盖所有北美DSRCT患者的36%。结论我们的数据表明,复发性EWSR1-WT1融合的连接是自然处理的,并在DSRCT细胞普遍存在的HLA等位基因的背景下进行了预处理。这些发现确定了融合派生的公共NEOAG是融合驱动的恶性肿瘤的可行的治疗靶标的来源。这项工作为针对EWSR1-WT1和其他复发性致癌融合的新型T细胞基于T细胞的疗法的临床翻译奠定了基础。

脊全瘤免疫的多模式分析揭示了不同的免疫情境
抽象的背景脊架是轴向骨骼中罕见的癌症,由于其解剖位置,其治疗选择有限,具有挑战性的临床管理。近年来,一些临床试验表明,丘多马斯可以对免疫疗法做出反应。然而,仍然缺乏对脊全瘤免疫力及其与临床参数的关联的深入描绘。方法我们通过应用多模式方法提出了76个脊架的免疫学特征的全面表征。进行了20个脊架的转录组分析,以通过抑制的免疫学常数(ICR)特征来告知免疫相关基因的活性。通过成像质量细胞仪进行多维免疫表型,以在32个脊架的不同免疫情境中提供见解。T细胞浸润,然后通过单变量和多元COX比例危害模型以及Kaplan-Meier估计值与临床参数相关。此外,在所有76名患者中,通过免疫组织化学染色评估了人类白细胞抗原(HLA)I类的不同表达模式。最后,通过对24名患者的TCRB基因座的可变区域进行分析,寻求T细胞受体(TCR)的克隆富集。结果通常表现出一种免疫“热”微环境,如ICR转录特征所示。此外,我们观察到大多数脊全瘤保持了HLA I类表达。我们根据T细胞浸润确定了两个不同的脊髓瘤,它们独立于临床参数。高度浸润的组进一步以较高的树突状细胞浸润和肿瘤中多细胞免疫聚集体的存在为特征,而低T细胞浸润与免疫和基质细胞的整体细胞密度较低有关。有趣的是,与TCR库的TCR库相比,TCR库的克隆富集更为明显。结论我们的发现揭示了通过识别

治疗性T细胞的基因组编辑
临床试验已证明,工程化的 TCR 𝛼𝛽 T 细胞具有作为白血病清除的有效介质的潜力,并且已授权的疗法正在用于治疗 B 细胞恶性肿瘤。虽然大多数应用都依赖于自体淋巴细胞的采集和操作,但基因组编辑技术的新兴应用已证明,同种异体 TCR 𝛼𝛽 细胞可以进行工程化以克服人类白细胞抗原 (HLA) 障碍,并提供了一种更具成本效益且广泛可及的“现成”疗法的途径。基因组编辑还提供了解决其他障碍(例如共享抗原表达)的前景,并且已应用于指导位点特异性转基因整合,以改善转录调控和功能。