XiaoMi-AI文件搜索系统
World File Search SystemUTR

USP mRNA 疫苗章节
几十年来,人们一直在探索利用信使核糖核酸 (mRNA) 技术来研发流感、寨卡病毒、狂犬病和巨细胞病毒等传染病的疫苗。COVID-19 疫情加速了该技术作为疫苗平台的研究和开发,导致 mRNA 疫苗成为美国首个获得紧急使用授权并随后获批用于 SARS-CoV-2 的疫苗。用于预防 COVID-19 的 mRNA 疫苗已被证明是该技术的成功应用,然而,对于检测这些疫苗质量属性的指导仍然有限。一套标准的分析方法将为世界各地的疫苗开发商、制造商、监管机构和国家控制实验室提供支持,通过提供工具来帮助加速使用该平台开发安全有效的疫苗,并防止出现劣质和伪造的疫苗产品。根据各利益相关方确定的这一需求,USP 和我们的 BIO3 专家委员会制定了 mRNA 疫苗的通用章节草案,作为制定 mRNA 疫苗测试程序章节的第一步。本章节包括分析程序和最佳实践,以支持对 mRNA 疫苗的共同质量属性进行评估。本章节草案还以一般章节<1235>《人用疫苗——一般考虑因素》和<1239>《人用疫苗——病毒疫苗》中描述的最佳实践为基础。章节草案中的方法改编自公开来源,尚未经过 USP 的核实或确认。USP 和我们的 BIO3 专家委员会将提前发布章节草案以征求公众意见。通过提前发布,USP 希望征求利益相关者对参考文件中描述的方法的反馈,并鼓励提交与章节草案中提出的方法相关的任何替代方法和任何其他支持文件,包括验证文件。引言天然存在的 mRNA 是在真核细胞中通过 RNA 聚合酶转录细胞核中的 DNA 来产生的。 mRNA 分子从细胞核运输到细胞质,在那里它们作为模板,由核糖体翻译产生特定的蛋白质。通过这种方式,储存在细胞核中的信息被用来产生特定的蛋白质。这种 mRNA 不能产生除其编码的蛋白质以外的任何蛋白质。注射后,mRNA 的估计半衰期约为 8-10 小时,之后它会迅速降解并被体内的天然 RNase 分解。mRNA 不需要进入细胞核即可发挥作用。通常,可以通过在宿主(例如大肠杆菌)中扩增起始 DNA 质粒来制备 mRNA 疫苗药物物质。质粒在用于大规模生产 mRNA 中间体之前,需要进行酶线性化和纯化。在无细胞系统中,通过体外转录从线性化质粒 DNA 模板中产生 mRNA。根据具体的制造工艺,构建体用核苷优化以形成序列,转录的 mRNA 在 7-甲基鸟苷的 5' 端酶促加帽和/或在 3' 端用 poly (A) 酶促加尾。然后纯化 mRNA 药物物质并配制成药物产品。mRNA 疫苗药物产品可以是嵌入脂质纳米颗粒 (LNP) 中的 mRNA 制剂。LNP 保护 mRNA 免于降解并帮助 mRNA 通过内吞作用进入细胞。一旦进入内体,mRNA 疫苗分子就会逃离内体进入细胞质(取决于可电离脂质和 mRNA 核苷酸的摩尔比)并提供模板以产生多个病毒蛋白拷贝。病毒蛋白作为抗原刺激免疫反应,这是疫苗接种的预期结果。目前已开发出两种主要形式的 mRNA 疫苗:非复制型 mRNA 疫苗(常规)和自扩增型 mRNA (SAM) 疫苗,如下图 1 所示。常规非复制型 mRNA 疫苗构建体通常由 5′ 7-甲基鸟苷帽结构、5′ 非翻译区 (UTR)、编码蛋白质的开放阅读框 (ORF)、3′ UTR 和 3′ poly(A) 尾组成。SAM mRNA 疫苗源自 alpha 病毒基因组,其中 mRNA 分子编码可指导细胞内 mRNA 扩增的其他复制酶成分。在这两种形式的 mRNA 疫苗中,UTR 区域对于最大化蛋白质表达、mRNA 分子的 5′ 加帽、阻断核酸外切酶介导的降解和提高翻译效率都很重要。UTR、5' 帽和 poly(A) 尾也有助于稳定非复制型 mRNA 疫苗(常规)和自扩增型 mRNA (SAM) 疫苗,如下图 1 所示。常规非复制型 mRNA 疫苗构建体通常由 5′ 7-甲基鸟苷帽结构、5′ 非翻译区 (UTR)、编码蛋白质的开放阅读框 (ORF)、3′ UTR 和 3′ poly(A) 尾组成。SAM mRNA 疫苗源自 alpha 病毒基因组,其中 mRNA 分子编码可指导细胞内 mRNA 扩增的其他复制酶成分。在这两种形式的 mRNA 疫苗中,UTR 区域对于最大化蛋白质表达、mRNA 分子的 5′ 加帽、阻断核酸外切酶介导的降解和提高翻译效率都很重要。UTR、5' 帽和 poly(A) 尾也有助于稳定非复制型 mRNA 疫苗(常规)和自扩增型 mRNA (SAM) 疫苗,如下图 1 所示。常规非复制型 mRNA 疫苗构建体通常由 5′ 7-甲基鸟苷帽结构、5′ 非翻译区 (UTR)、编码蛋白质的开放阅读框 (ORF)、3′ UTR 和 3′ poly(A) 尾组成。SAM mRNA 疫苗源自 alpha 病毒基因组,其中 mRNA 分子编码可指导细胞内 mRNA 扩增的其他复制酶成分。在这两种形式的 mRNA 疫苗中,UTR 区域对于最大化蛋白质表达、mRNA 分子的 5′ 加帽、阻断核酸外切酶介导的降解和提高翻译效率都很重要。UTR、5' 帽和 poly(A) 尾也有助于稳定
基于 CRISPR 的基因编辑增强 LDLR 表达...
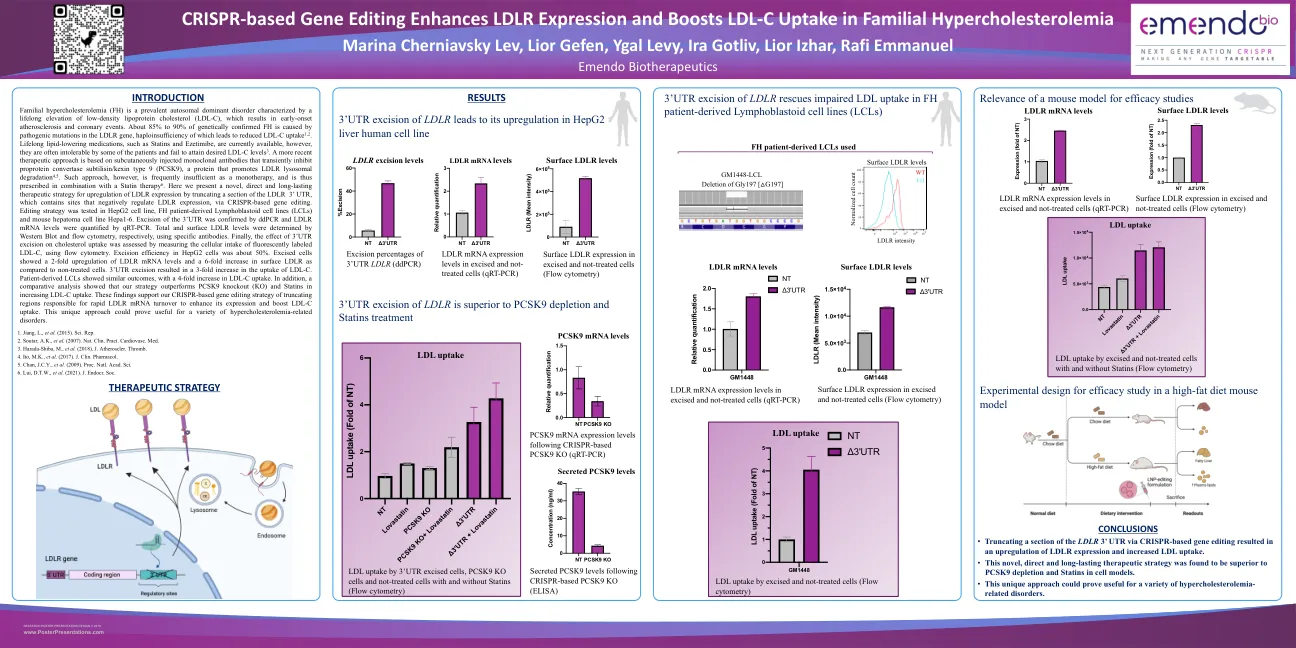
家族性高胆固醇血症 (FH) 是一种常见的常染色体显性遗传病,其特征是低密度脂蛋白胆固醇 (LDL-C) 终生升高,导致早发性动脉粥样硬化和冠状动脉事件。约 85% 至 90% 经基因确诊的 FH 是由 LDLR 基因致病突变引起的,该基因的单倍体不足会导致 LDL-C 摄取降低 1,2 。目前可以使用他汀类药物和依折麦布等终生降脂药物,但有些患者往往无法耐受这些药物,无法达到理想的 LDL-C 水平 3 。一种较新的治疗方法是基于皮下注射单克隆抗体,这种抗体可以暂时抑制前蛋白转化酶枯草溶菌素/kexin 9 型 (PCSK9),PCSK9 是一种促进 LDLR 溶酶体降解的蛋白质 4,5 。但这种方法作为单一疗法往往是不够的,因此需要与他汀类药物联合使用 6 。在这里,我们提出了一种新颖、直接且持久的治疗策略,通过基于 CRISPR 的基因编辑截断 LDLR 3' UTR 的一部分(其中包含负向调节 LDLR 表达的位点),以上调 LDLR 表达。在 HepG2 细胞系、FH 患者来源的淋巴母细胞系 (LCL) 和小鼠肝癌细胞系 Hepa1-6 中测试了编辑策略。通过 ddPCR 确认 3'UTR 的切除,并通过 qRT-PCR 量化 LDLR mRNA 水平。分别通过 Western Blot 和流式细胞术使用特异性抗体测定总 LDLR 水平和表面 LDLR 水平。最后,通过流式细胞术测量荧光标记的 LDL-C 的细胞摄入量来评估 3'UTR 切除对胆固醇摄取的影响。 HepG2 细胞中的切除效率约为 50%。与未经处理的细胞相比,切除的细胞显示 LDLR mRNA 水平上调 2 倍,表面 LDLR 增加 6 倍。3'UTR 切除导致 LDL-C 摄取增加 3 倍。患者来源的 LCL 显示出类似的结果,LDL-C 摄取增加 4 倍。此外,比较分析表明,我们的策略在增加 LDL-C 摄取方面优于 PCSK9 敲除 (KO) 和他汀类药物。这些发现支持我们基于 CRISPR 的基因编辑策略,即截断负责快速 LDLR mRNA 周转的区域以增强其表达并促进 LDL-C 摄取。这种独特的方法可能对多种高胆固醇血症相关疾病有用。

遗传变异调节炎症体活性
在炎症体启动和激活后,对骨row衍生的巨噬细胞(BMDM)进行了白细胞介素-1 B释放的抽象定量性状基因座映射(BMDM)。最强的相关基因座映射到7号染色体上的Pycard基因,该基因座的染色体基因编码为炎症体适配器蛋白凋亡相关的斑点样蛋白(ASC)(ASC)。DBA/2和AKR Pycard基因在其3'未翻译区域(UTR)中仅在单核苷酸多态性(SNP)上有所不同。DBA/2与AKR BMDM的pycard mRNA表达和ASC蛋白水平升高,并增加了炎性体斑点的形成,这与Pycard mRNA稳定性的增加有关,而转录率没有提高。crispr/cas9基因编辑在DBA/2胚胎干细胞上进行,以将pycard 3'utr SNP从dba/2更改为AKR等位基因。由于pycard mRNA稳定性降低,将细胞分化为巨噬细胞后,这种单碱基的变化显着降低了Pycard表达和炎症体活性。

mRNA:向医学传递信息
• 细胞质多聚腺苷酸化元件结合蛋白 (CPEB1) • 与 3'UTR、CPSF 中富含 A/U 的 CPE 相互作用 • PARN(多聚腺苷酸核酸酶)和 Gld2(多聚腺苷酸聚合酶)的酶活性相反 • Aurora A 对 CPEB1 的磷酸化释放 PARN,Gld2 活性占主导地位 • PABPC1 关联、多核糖体(帽子相互作用)和翻译激活

TrkB.T1 作为慢性疼痛的治疗靶点
原肌球蛋白相关受体激酶 B (TrkB) 是脑源性神经营养因子 (BDNF) 的受体;其信号传导通过激活几个下游级联,有助于神经元存活、可塑性、分化和生长。缺乏细胞内激酶结构域的截短异构体 (TrkB.T1) 的过度表达与慢性疼痛的发展和持续有关。已发表的数据显示,小鼠模型中的 TrkB.T1 敲除可恢复运动功能并减轻脊髓损伤后的疼痛。我们项目的目标是确定抑制 TrkB.T1 表达的小分子作为慢性疼痛的潜在疗法,重点关注两种调节机制:(1) TrkB 前 mRNA 的差异转录后加工,以及 (2) 通过其 mRNA 的 3' 非翻译区 (3'UTR) 对 TrkB.T1 表达的转录后调节。对于第一点,我们假设两种主要 TrkB 亚型的比例主要受上游 (T1) pA 位点的切割和多聚腺苷酸化 (pA) 位点识别控制,因此抑制该位点 3'-加工的药物应能抑制 TrkB.T1 合成。对于第二点,我们假设 TrkB.T1 mRNA 3'UTR 包含调节序列,这些序列的功能可通过操纵关键反式因子的功能、表达或 RNA 结合活性的化合物进行调节,从而通过加速 TrkB.T1 mRNA 的衰变和/或抑制其翻译来抑制 TrkB.T1 的产生。对于每种机制,我们开发了独立的活细胞高通量筛选 (HTS) 检测方法,以识别可以 (1) 阻断 TrkB.T1 pA 位点的 3'-切割和多聚腺苷酸化,或 (2) 通过 TrkB.T1 mRNA 3'UTR 抑制基因表达的小分子。利用这些发现,我们旨在发现一种或多种能够抑制 TrKB.T1 表达的新药物,这些药物可在慢性疼痛的小鼠模型中作为新型镇痛药进行测试。

补充表1。siRNAS序列
PCDNA3.1 >NM_001419.3: 164-1144 Homo sapiens ELAV like RNA binding proteins 1 (ELAVL1), mRNA, GCCCATGTCTS GICKECICIPHASIBILITIES GATCGTCACTCCTCCTCCTITIES CAGGAGGAS TGOGTGTCITCATGTGTGIGHTGTGITS TGAATCTS ttcgghtstcghts acacagtgtgtgtgtgtghtghtghtghtghtghtghtghtghtgiciciescscggticugis cgcgiaghgiciphatgiciphaciciphaciciphaciciphasivaities casciaciaciagiagiaties cccagaggicipegicesgttcgtcgtgtgtgtgtcctcggtcgtcgtcgtcgtcgtcgtcgtcgtcgtcgtgtgtgtgtgtgtgtscicciccicciccciccgginigiro agtttttttttttttcaatgtgtgtgtgtgtgtgtcccagtcctgtgag CCCATCAGTGTTGTGTGCAGCACCACCACCACCACCACCACCACCACCICCICCCICCICCIachIs CACCAGCGCAGATTCAGTCTCCATGGC GTCTCATGEGCTGTCTGTCGTCGTGTGTGTGTGTGTGTGTCCTCCTCCTITCTCTCITCATCSCIOUS ctgcagatgtggggtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgitgitgitgitghacipecthingshings gcaagtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgitgitgitgitgitgitgittgitgittgitsgittsgitsgittsssss。 taccgcctggggggggggggggggggggggggggggggggggggggttctcs aaaaccaagtcccccccccccccccccccccccccedccccccccccccadcccccccsccadcccccccccccadcccccccccccccadcccccccccccsccadcccccccccccadccccccccccccccadcccccccccccscc-ad一下,,

labgscan fraxa套件
FMR1基因编码称为FMRP的RNA结合蛋白(脆弱的X智力低下蛋白),位于XQ27.3中,具有17个外显子,延伸超过39 kb的基因组DNA。该基因在5'UTR区域具有CGG多态性重复。此重复序列中的突变是大多数情况的原因:

上电力传输有限公司招标规范编号...
没有电子招标文件费、文件购买费和 EMD 的公司投标将不予开标。电子招标文件费将通过 RTGS/NEFT 存入上述 UPPTCL 账户,并提供此类存款证明,例如UTR 号、账户名称、投标人签字的存款单扫描件应与电子招标文件一起上传。保证金 (EMD) 将通过 RTGS/NEFT 存入上述 UPPTCL 账户,并提供此类存款证明,例如投标人应与 EMD 一起上传 UTR 号、账户名称、投标人签字的存款单扫描件。保证金 (EMD) 将通过 RTGS/NEFT 存入上述 UPPTCL 账户,并提供此类存款证明,例如UTR 号、账户名称、投标人副署的付款单扫描件应与电子招标文件一起上传,或以银行担保的形式上传给电力 765/400kV 变电站设计环 II、UPPTCL、勒克瑙的监理工程师。如果 EMD 以银行担保的形式存入,则 EMD B.G. 的扫描件。以及开具 B.G. 的银行发出的确认邮件的扫描件。将与电子招标文件一起上传。投标人必须在电子投标第一部分开始后 7(七)天内或电子投标第二部分开始前(以较早时间为准)向电力 765/400kV 变电站设计圈 II 的监理工程师办公室提交同一份银行担保的原件。如果未提交 EMD 银行担保原件,则投标人提交的电子投标第二部分将不予开标。无需以纸质或信使形式提交包括投标费、EMD B.G.和其他商业文件的招标文件。包含价格投标的第二部分将在稍后单独开标,并应相应通知投标人

利用纳米技术针对病毒基因组的结构特征......
对于大多数病毒来说,这种识别所需的信息目前还没有,而且与 RNA 结构结合的药物晶体结构很少(并且不一定具有代表性);迫切需要在分子水平上重新认识这种结合。RNA 分子固有的灵活性使 RNA 结构研究变得更加复杂,这需要了解它们的动力学而不仅仅是它们的基态构象。因此,简单的分子对接是不够的;相反,分子动力学可以潜在地探测能量图和结构灵活性。在这里,我们首次采用分子动力学详细探索纳米级药物插入病毒 RNA UTR 凸起部分,复制实验观察结果并对 RNA 动力学和药物进入过程获得全新的认识;这为设计新型 UTR 结构靶向药物提供了重要信息。所研究的纳米级药物是超分子圆柱体,它不仅具有前所未有的 RNA 凸起结合能力,而且是金属超分子结构中第一个在细胞测定中表现出强效抗病毒活性的药物。35 人们对金属超分子结构在生物学中的应用越来越感兴趣。36 – 41

NDP 外显子 2 区域 KO 报告模板...
图 S01:CRISPR/Cas9 编辑人类 iPSC。(A) 使用双 sgRNA(sg1 和 sg2)靶向外显子 2 两侧的区域来生成 HDR 介导的 NDP KO-GFP 报告系,使用带有 KO 报告模板的 PUC57 载体,由外显子 2 (5'UTR)-eGFP-P2A 盒组成,两侧是同源臂 (HA),其中包括 PAM 位点突变和用于质粒线性化的 sgRNA 切割位点 (sg1-c 和 sg2-c)。在此过程中还生成了 NHEJ 介导的 NDP KO (NDP Δ exon2) 以及 NDP WT 同源克隆。(B) 使用类似方法生成 NDP WT-GFP 报告系。外显子 3 的 CDS 两侧的双 sgRNA(sg3 和 sg4)和含有修复模板的质粒,由外显子 3(CDS)-V5-P2A- eGFP 盒组成,两侧是同源臂,包括 PAM 位点突变和用于质粒线性化的 sgRNA 切割位点(sg3-c 和 sg4-c)。sgRNA,小向导 RNA;HDR,同源定向修复;KO-GFP,敲除报告基因;KO,敲除;WT,野生型;PAM,protoscpacer 相邻基序;UTR,非翻译区;CDS,编码序列;HA,同源臂;eGFP,增强型绿色荧光蛋白;P2A,猪 teschovirus-1 2A 自切割肽;V5,V5 标签。