XiaoMi-AI文件搜索系统
World File Search Systemautism

SA 自闭症战略 2024-2029 咨询
2. AMAZE,《澳大利亚人对自闭症的态度和行为》,2018 年 3. NDIS,《自闭症摘要仪表板》,2023 年 4. 国家残疾保险机构,《国家残疾保险计划 (NDIS)》季度报告 2022-2023 年。 5. ABS,《残疾、老龄化和护理人员调查》,2019 年 6. Amaze,《谈论自闭症:尊重和准确报道自闭症和自闭症患者的指南》,2019 年 7. Amaze,《谈论自闭症:尊重和准确报道自闭症和自闭症患者的指南》,2019 年 8. Amaze,《澳大利亚的自闭症和就业》,2019 年 9. ABS,《残疾、老龄化和护理人员调查》,2019 年 10. NDIS,《自闭症摘要仪表板》,2023 年 11. NDIS,《自闭症摘要仪表板》,2023 年


维多利亚时代的自闭症计划2023刷新
我们深深致力于原住民的自决,并支持维多利亚的条约和真相的过程。我们承认,条约将对我们与原住民维多利亚时代的工作产生广泛的影响,我们致力于通过维多利亚时代的自闭症计划采取调查过程中的发现。我们寻求建立尊重和协作的伙伴关系,并制定尊重原住民自决并与条约愿望保持一致的政策和计划。

自闭症,患病率和与童年的联系激增...
抽象自闭症一种神经退行性障碍,其特征是儿童行为异常,诸如孤立,学习困难,沟通和社会互惠的普遍异常,无法保持眼神接触或对某事,与周围视力和许多其他神经系统疾病的事物以及许多其他神经系统疾病,包括自闭症的社交因素,Epig extical,Epig ecig ancig ecig ancig ecig ancig ancig ancig ecig ancig ancig ecig ancig and Epig ecig ancig ecig and epig,疫苗在自闭症中发挥了主张的作用,这引起了疫苗的极大兴趣,这主要是由于许多疫苗中含有复合硫镁的汞含量较高,并且由于一些证据与某些疫苗联系起来,这些证据将包括麻疹,腮腺炎和红宝石疫苗在内的某些自闭症与自闭症相关的疫苗联系起来,而不是几种真实的科学研究。在这里,我们批判性地回顾了这些断言,检查了疫苗的一些含义成分,并将其与自闭症的神经生物学联系起来。关键词:自闭症,神经病理学,硫柳汞,疫苗接种,儿童


国家自闭症策略概述-Amaze
我们缺乏针对优先人群的特定需求并解决交叉劣势的承诺感到失望。我们还担心该策略包含一些显着差距。例如,它不包括与住房有关的承诺,也没有关于教育,正义,父母和看护者的承诺。它没有致力于建立特定自闭症的倡导服务,也只专注于澳大利亚政府,而不是各州和地区。我们认为国家一致性很重要。
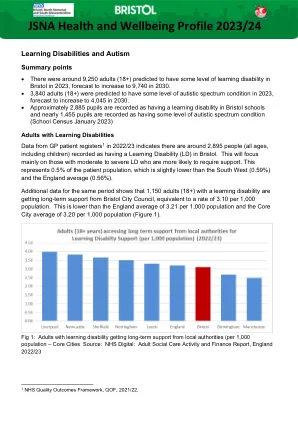
JSNA 2023.24-学习障碍和自闭症
学习障碍的人的健康状况比一般人群差;很多都是可以避免的。在需要时获得有效医疗保健的困难会使它们变得更糟4。国家研究5几年前的5年前,学习障碍的男性平均死亡比更广泛的人口和女性早于20年的时间早13年,并且对学习障碍患者的健康状况率提高,包括癫痫,心理健康和心脏病。自闭症频谱条件

催产素,gaba和自闭症中的多巴胺相互作用
催产素在大脑发育中起重要作用,并且与大脑中的各种Neu Rotransitter系统有关。至少在发育的某些阶段,催产素在大脑中产生,分泌和分布的异常对于神经精神疾病的病原体至关重要,尤其是在自闭症谱系疾病中。自闭症的病因包括大脑的局部感觉和多巴胺能区域的变化,这也由催产素的下丘脑来源提供。了解他们的相互关系非常重要。在本综述中,讨论了催产素与多巴胺耐药系统,γ-氨基丁酸(GABA)抑制性神经传递及其在自闭症谱系障碍中的变性的关系。特别关注的结果描述了大脑抑制性GABA能标记的表达降低。据推测,由于催产素在某些发育阶段缺乏或功能障碍,GABA能Neu Rotransersission会改变,因此抑制了多巴胺能信号传导并有助于自闭症症状。

自闭症患者的数字包含
实际意义:研究中考虑的隐藏或潜在残疾的各个方面为就业提供了不同的观点,重点是可以创造的工作场所条件。雇主可能不知道需要为那些隐藏健康问题的人创建特殊的工作条件。这会对失业,病假增加,工作场所的机会有限等产生负面影响。员工通常不愿向雇主披露其非明显健康问题;因此,雇主应通过创建相关条件来促进披露此类信息。这种方法将有助于对这类员工的法律保护,并进一步发展现有的立法法规,因为后者不完全遵守当今的需求并改变了现实。

自闭症儿童与自闭症儿童的处理速度比较
本研究比较了自闭症谱系障碍 (ASD) 儿童和智力障碍 (ID) 儿童的处理速度,以研究这两种发育状况之间的认知差异。处理速度是一项关键的认知功能,对学业成绩和日常功能至关重要,但自闭症和智力障碍儿童的处理速度往往受到损害。研究结果表明,智力障碍儿童的处理速度比自闭症同龄人要慢。这些差异可能归因于潜在的认知和神经发育因素,例如工作记忆、注意力和执行功能的缺陷,这些缺陷在智力障碍儿童身上更为明显。此外,该研究还强调了智力障碍儿童的处理速度缺陷如何对学业和社会结果产生负面影响。