XiaoMi-AI文件搜索系统
World File Search SystemcDNA

使用高灵敏纤维微腔生物传感器
摘要:开发了一种基于微腔纤维马赫德 - Zhhnder干涉仪的新型无标签光纤生物传感器,并实际上证明了用于DNA检测的。使用偏置剪接标准通信单模纤维(SMF)制造生物传感器。传感器的光路径受偏置开放腔中液体样品的影响。在实验中,在折射率(RI)测量中实现了-17,905 nm/riU的高灵敏度。在此基础上,探针DNA(pDNA)使用APTES固定在传感器表面上,从而实现了捕获的互补DNA(cDNA)样品的实时监测。实验结果表明,生物传感器的高灵敏度为0.32 nm/fm,检测限为48.9 AM。同时,传感器具有高度可重复和特定的性能。这项工作报告了易于制造,超敏感和无标签的DNA生物传感器,该生物传感器在医学诊断,生物工程,基因识别,环境科学和其他生物领域中具有重要的潜在应用。

luxturna®(voreTigene neparvovec-rzyl)
遗传性视网膜营养不良是由基因突变引起的一组眼睛疾病。RPE65基因以操作顺序以保持光感应感光受体细胞,视网膜的棒和锥所需的一种维生素A。RPE65基因突变会导致RPE65蛋白质功能的部分或总损失,这可能会导致严重的视力障碍,从生命的早期开始。用于治疗RFE65突变相关的视网膜疾病的基因治疗使用视网膜下注射病毒载体,将治疗基因插入具有已知RPE65突变的人的可行视网膜细胞中。正常等位基因的引入可以纠正现有的可行视网膜细胞中的突变基因,并将这些细胞返回正常功能。腺相关病毒(AAV)是最安全,最有效的病毒载体。voretigene Neparvovec(Luxturna)是含有腺相关的病毒2(AAV2)载体,含有人RPE65互补DNA(CDNA),迄今为止,是FDA批准的基因疗法,用于治疗RPE65遗传疾病的治疗。

ARID1A参与胃癌的DNA双链断裂修复
使用Trizol试剂(Invitrogen,Carlsbad,CA,美国)提取总RNA。使用Prime-Script RT试剂试剂盒(完美的实时,日本Takara)合成互补的DNA(cDNA)。3-磷酸甘油醛脱氢酶(GAPDH)用作内部对照基因,并使用2 – DDCT公式计算折叠诱导。使用SYBR Premix ex Taq进行定量实时PCR分析(日本Takara。Inc.目录编号DRR041A)(PCR协议:阶段1:早期讲述,重复:1;95℃30s阶段2:PCR反应,重复:40; 95℃5 s; 60℃30s阶段3:熔体曲线:熔体曲线:95℃15s; 60 s; 60 s; 60 s; 95 s; 95 s; 95 s;95℃15s)。使用了以下引物:ARID1A(F)5'-CTTCACTCAGTCAGCTCCCA-3',arid1a(r)5'-GGTCACCCCACCCTCATCTCATACTCCTTT-3',gapdh(f)5'-GGGTGGTGGTGGTGGTGGTGGTGGTGGTCTCTCTCTCTCTCCTGATCTCAACA-3',and gapdh(R) 5'-GTTGCTGCCCAAATTCGTTGT-3'。GAPDH用作内源性对照。每次三份重复每个样本,并使用相对定量软件(Applied Biosystems)分析。

编码淋巴细胞寄托受体CD44的基因组结构至少揭示了12个剪接外显子
摘要CD44分子已知表现出大小的异质性,这既归因于替代剪接和差异糖基在繁殖域内的差异糖基化。尽管从cDNA测序中部分推断出了几个替代外显子的存在,但据我们所知,尚未描述CD44基因的精确内含子外观。在本研究中,我们描述了人类CD44基因的结构,该基因至少包含19个外显子DNA的大约50个基因酶。我们已经确定了10个在细胞外域内的剪接外显子,包括1个以前没有报道的外显子。除了整个外显子的cluson或辩解外,还通过在单个外显子2中的内部剪接供体和受体位点的uztion产生更多的多样性。先前报道的细胞质结构域的变异表明是由2个外显子的替代剪接引起的。CD44的基因组结构揭示了显着的复杂性,我们证实了替代剪接作为CD44分子中结构和功能多样性的基础的作用。

一个细胞竞争系统,具有一个基因表达的单拷贝基因中的一个基因
即使使用晚期质粒和病毒载体,在不同转染的细胞中获得多个基因的拷贝数也很具有挑战性。,我们使用转基因竞争系统,Kazusa cDNA克隆和我们的双重重组酶介导的盒式盒式交换系统的组合从一个细胞中的单拷贝基因中实现了一个基因表达。使用该系统在HEK293中同时在同一表达水平中同时表达了所有48个核受体,并比较了细胞增殖速率。在8周后用CMV-或EF1启动子驱动子驱动的表达转染的细胞之间观察到显着差异。EF1-NR1I2细胞系从2周到8周增长,比EF1-DSRED线高1.13倍。另一方面,EF1-NR4A1细胞系在8周时显示最大减少,比EF1-DSRED线低0.88倍。在我们的转基因竞争系统和长期增长实验中证实了结果。我们的转基因竞争系统提供了广泛,简单且准确的细胞竞争方法。
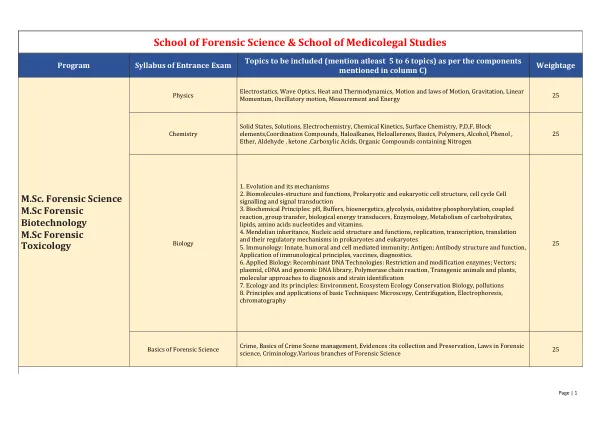
NTA 入学考试大纲 (1).pdf
1. 进化及其机制 2. 生物分子的结构和功能、原核和真核细胞结构、细胞周期、细胞信号传导和信号转导 3. 生化原理:pH、缓冲液、生物能学、糖酵解、氧化磷酸化、偶联反应、基团转移、生物能量转换器、酶学、碳水化合物、脂质、氨基酸核苷酸和维生素的代谢。 4. 孟德尔遗传、核酸的结构和功能、原核生物和真核生物的复制、转录、翻译及其调控机制 5. 免疫学:先天、体液和细胞介导免疫;抗原;抗体的结构和功能、免疫学原理的应用、疫苗、诊断学。 6. 应用生物学:重组 DNA 技术:限制和修饰酶;载体;质粒、cDNA和基因组DNA文库、聚合酶链反应、转基因动物和植物、分子诊断和菌株鉴定方法7.生态学及其原理:环境、生态系统生态学保护生物学、污染8.基本技术的原理和应用:显微镜、离心、电泳、色谱法

dmt改变皮层行进波
抽象的许多生物在自然界中具有进化的机制,可以耐受严重的低氧或缺血,包括具有冬眠能力的北极松鼠(AGS)。尽管AGS中的低氧或缺血耐受性涉及生理适应性,但对内在AGS细胞弹性对代谢应激的弹性的关键细胞机制知之甚少。通过基于细胞存活的cDNA表达筛选在神经祖细胞中,我们确定了AGS ATP5G1的遗传变异,该变异赋予了细胞对代谢应激的弹性。ATP5G1编码线粒体ATP合酶的亚基。在小鼠细胞中的异位表达和内源性AGS基因座的CRISPR/CAS9基础编辑表明,一个AGS特异性氨基酸取代在介导AGS ATP5G1的细胞保护过程中的因果作用。AGS ATP5G1通过调节线粒体形态变化和代谢功能来促进代谢应激的弹性。我们的结果从哺乳动物的冬眠器中确定了ATP5G1的自然发生的变体,该变体对代谢应激有助于固有的细胞保护作用。

图S1。 GADD45β和...
BNT162B2尖峰mRNA定量分析。用于定量BNT162B2 SPIKE mRNA的相对含量,如Gasparello等人(1)所述,在24小时处理后分离了来自BNT162B2处理的细胞的总RNA,并将其反转录为CDNA。Fertig等人(2)对BNT162B2 SPIKE mRNA的定量(Q)PCR分析进行了。在这项研究中,底漆被设计为特定于公共可用的密码子优化疫苗mRNA序列(2,3),但不是编码S蛋白的病毒RNA。使用的引物的序列为5'-GTG GAT CTG CCC ATC GGC ATC(正向)和5'-GTC CAT CCG CCG CTG CTG CTG CTA TCG(反向),并以200 nm的最终浓度使用。对于PCR分析,在95°C下首次10分钟后,进行了40个循环放大周期(在95°C时为15秒,在65°C时1分钟),并从60至95°C进行加热的最后一步。PCR扩增

引用:Abdullah M,Furtado A,Masouleh AK,Okemo P,Henry RJ。 2024年。改进的单倍型解析基因组揭示了更多的水稻基因。热带植物
Nipponbare是一种Japonica水稻品种,已被广泛用作水稻的标准参考基因型[1]。大米(Nipponbare)基因组是20多年前测序的最早测序的作物基因组之一[2]。大米基因组的第1个序列于2002年完成,是国际水稻基因组测序项目,2005年的植物基因组学领域的主要英里石[3]。这些国际合作努力提供了作物工厂的第一个基因组。Nipponbare基因组组装含有间隙,主要是由于重复的DNA序列。在2005年,这些差距总共约为18.1 MB,大部分来自centromeres和端粒区域。对技术进步和正在进行的研究工作的测序,随着时间的推移改善了水稻基因组序列[4,5]。thor的努力,以提高2013年的裸露参考基因组组件的质量,从而大大提高了cDNA序列和基因注释的精度,而它仍然不完整[5]。在人类基因组中,在组装和特征化方面已取得了最新的迈进,先前未开发的8%的人类基因组,尤其是包括端粒序列[6]。

硕士课程大纲
课程描述目录:教授先进的核酸修饰和分析方法。包括定点诱变、DNA 测序和 RNA 分析方法、用于基因分型的高分辨率 DNA 熔解和用于定量样本中 DNA 的实时 PCR。结合使用 CRISPR 基因编辑技术突变 2 个基因的方法,然后使用 RT-PCR 分析基因表达(RNA 分离、创建 cDNA,然后进行实时 PCR)。本课程教授基因工程和基因表达中使用的先进技术。这些包括体外诱变、DNA 测序、高分辨率 DNA 熔解、实时 PCR、RT-PCR 和 CRISPR 基因编辑的原理。您还应该学习科学方法、故障排除、数据分析以及如何独立思考和执行。课程属性 本课程具有以下属性: ☐ 通识教育要求 ☐ 全球/跨文化毕业要求 ☐ 写作强化毕业要求 ☒ 课程学科核心要求 ☐ 课程选修课核心要求 ☐ 开放选修课 其他:单击此处输入文本。 讲师信息 讲师姓名:生物技术教授 学生学习成果