XiaoMi-AI文件搜索系统
World File Search Systemcircuits

EE5M02 微电子电路
该模块将使学生深入了解电路和系统的超大规模集成 (VLSI)。该模块的最终目标是让学生掌握足够的知识,能够将大型数字电路的功能描述(硬件描述语言 (HDL) 级别)转换为物理布局描述(通常使用 GDSII 格式),适合在代工厂进行制造(流片)。该模块的结构分为两部分。VLSI 电路组件每周分配两次讲座,涵盖设备物理特性,重点关注非理想晶体管行为、电路和线路延迟模型、VLSI 电路复杂性的数学模型和产量估算。VLSI 系统组件每周分配一次讲座,涵盖用于实现电子设计自动化 (EDA) 流程的复杂软件工具链中使用的算法和数据格式。这两个实验室都基于 VLSI 系统讲座。

光子集成电路
北约中的科技组织 北约背景下的科学技术 (S&T) 被定义为为国防和安全目的有选择地和严格地生成和应用最先进的经过验证的知识。科技活动包括科学研究、技术开发、转化、应用和现场测试、实验和一系列相关的科学活动,包括系统工程、运筹学和分析、综合、集成和验证通过科学方法获得的知识。北约 STO 的使命是帮助将国家和北约的科技投资定位为北约国家及其伙伴国家国防和安全态势的知识和技术优势的战略推动者,方式如下: o 开展和促进科技活动,增强和利用联盟、北约国家和伙伴国家的能力和计划,支持北约的目标; o 按照北约的政策,增强北约在北约国家和伙伴国家中推动和影响安全和防御相关能力发展及威胁缓解的能力;o 支持北约国家和北约的决策。


ELEC2134电路和信号
课程协调员:Julien Epps教授,EE105室,j.epps@unsw.edu.au教程实验室协调员:Cameron.jones先生,Cameron.jones@unsw.edu.au咨询:您鼓励您在课程中或在课程中询问课程的材料,而不是在课程中,而不是通过电子邮件进行。 咨询时间将每周在讲座和Moodle上的建议时每周举行。 对于与教程相关的问题,最好将它们发布到讨论论坛中,但也欢迎您直接发送电子邮件。 所有电子邮件查询都应在主题行中的ELEC2134与UNSW学生的电子邮件地址进行,以确保将回答。 保持知情:可以通过电子邮件(您的学生电子邮件地址)和/或通过在线学习和教学平台在上课期间发布公告 - 在本课程中,我们将使用Moodle https://moodle.telt.unsw.edu.au/login/login/login/index.php。 请注意,您将被视为已收到以这种方式分发的所有信息,因此您应该仔细记下所有公告。课程协调员:Julien Epps教授,EE105室,j.epps@unsw.edu.au教程实验室协调员:Cameron.jones先生,Cameron.jones@unsw.edu.au咨询:您鼓励您在课程中或在课程中询问课程的材料,而不是在课程中,而不是通过电子邮件进行。咨询时间将每周在讲座和Moodle上的建议时每周举行。对于与教程相关的问题,最好将它们发布到讨论论坛中,但也欢迎您直接发送电子邮件。所有电子邮件查询都应在主题行中的ELEC2134与UNSW学生的电子邮件地址进行,以确保将回答。保持知情:可以通过电子邮件(您的学生电子邮件地址)和/或通过在线学习和教学平台在上课期间发布公告 - 在本课程中,我们将使用Moodle https://moodle.telt.unsw.edu.au/login/login/login/index.php。请注意,您将被视为已收到以这种方式分发的所有信息,因此您应该仔细记下所有公告。



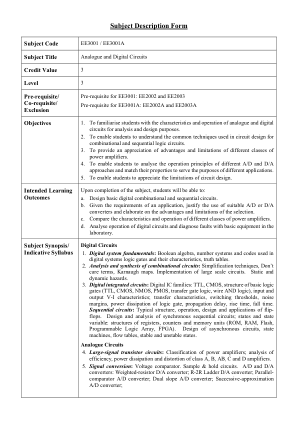
模拟和数字电路
1. 数字系统基础:布尔代数、数字系统中使用的数字系统和代码、逻辑门及其特性、真值表。2. 组合电路的分析与综合:简化技术、无关项、卡诺图。大规模电路的实现。静态和动态风险。3. 数字集成电路:数字 IC 系列:TTL、CMOS、基本逻辑门结构(TTL、CMOS、NMOS、PMOS、传输门逻辑、线与逻辑)、输入和输出 VI 特性;传输特性、开关阈值、噪声容限、逻辑门的功率耗散、传播延迟、上升时间、下降时间。时序电路:触发器的典型结构、操作、设计和应用。同步时序电路的设计和分析;状态和状态变量:寄存器、计数器和存储器单元(ROM、RAM、Flash、可编程逻辑阵列、FPGA)的结构。异步电路的设计、状态机、流表、稳定和非稳定状态。


代数 Bethe 电路
洛斯阿拉莫斯国家实验室是一家采取平权行动/提供平等机会的雇主,由 Triad National Security, LLC 为美国能源部国家核安全局运营,合同编号为 89233218CNA000001。通过批准本文,出版商承认美国政府保留非独占的、免版税许可,可以为了美国政府的目的出版或复制本文的已发表形式,或允许他人这样做。洛斯阿拉莫斯国家实验室要求出版商将本文注明为在美国能源部的支持下完成的工作。洛斯阿拉莫斯国家实验室坚决支持学术自由和研究人员的发表权利;但是,作为一个机构,实验室并不认可出版物的观点,也不保证其技术上的正确性。